

Activation of the Wnt/planar cell polarity pathway is required for pericyte recruitment during pulmonary angiogenesis
- PMID: 25447046
- PMCID: PMC4278244
- DOI: 10.1016/j.ajpath.2014.09.013
Activation of the Wnt/planar cell polarity pathway is required for pericyte recruitment during pulmonary angiogenesis
Abstract
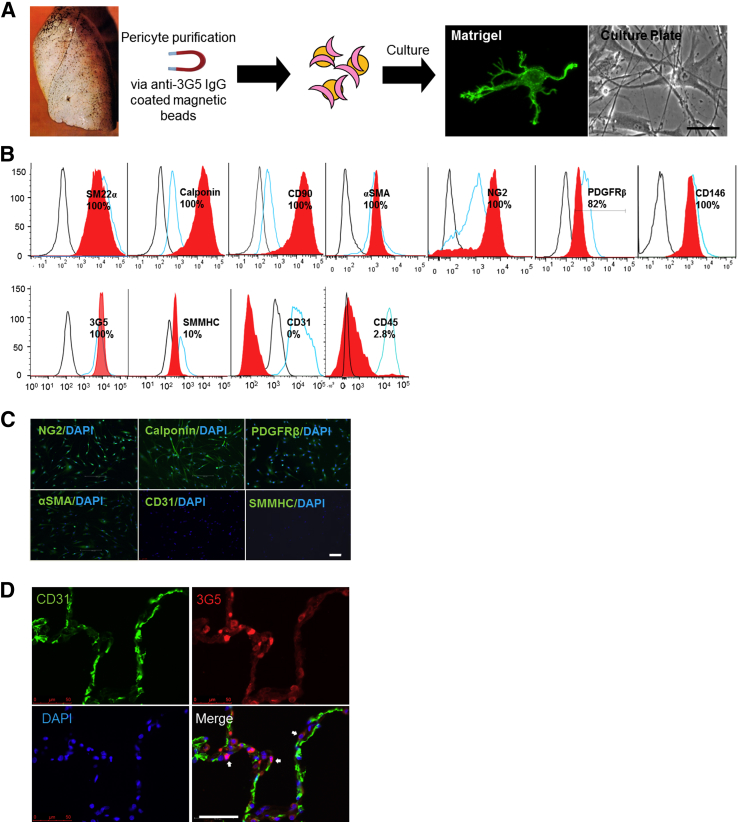
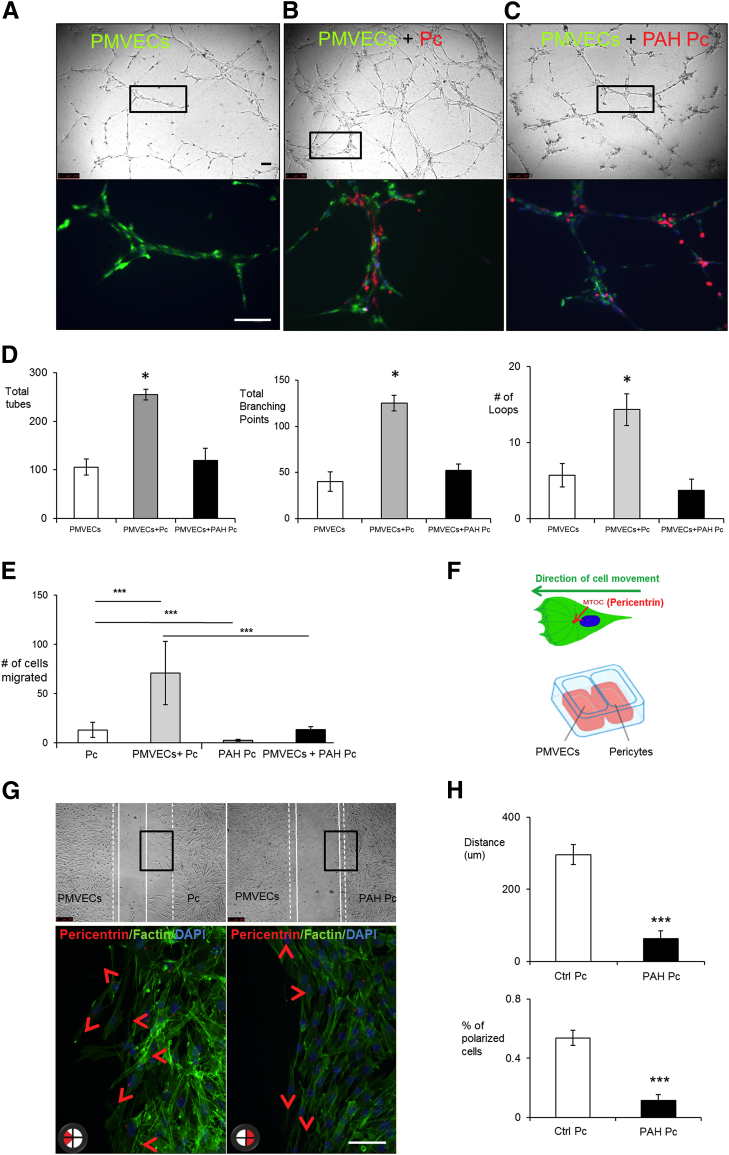
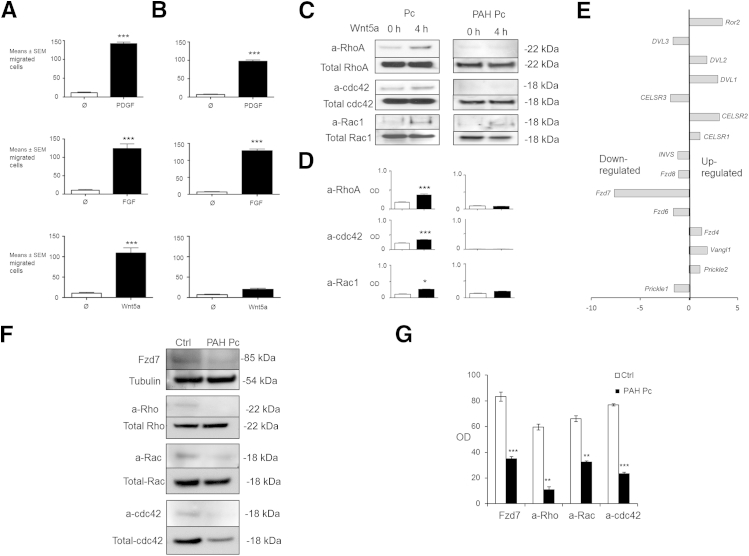
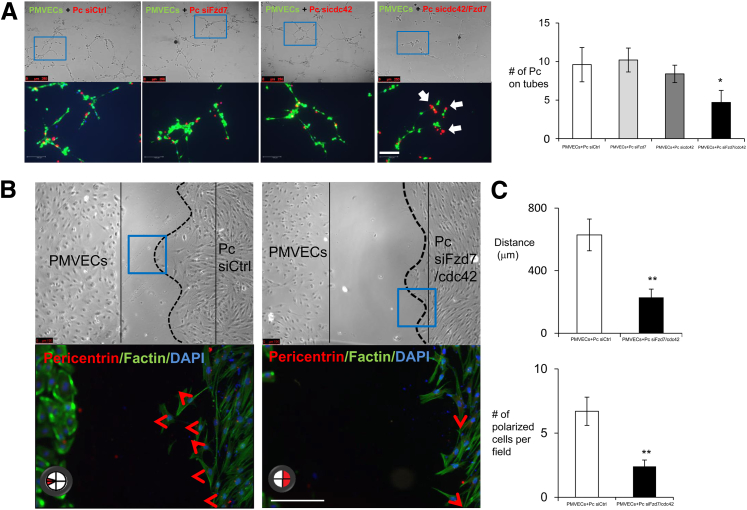
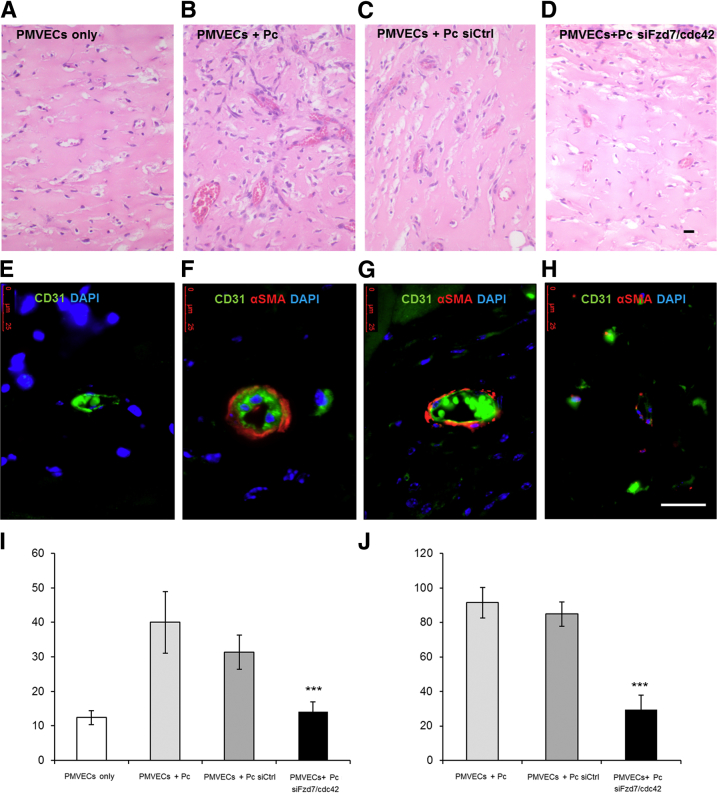
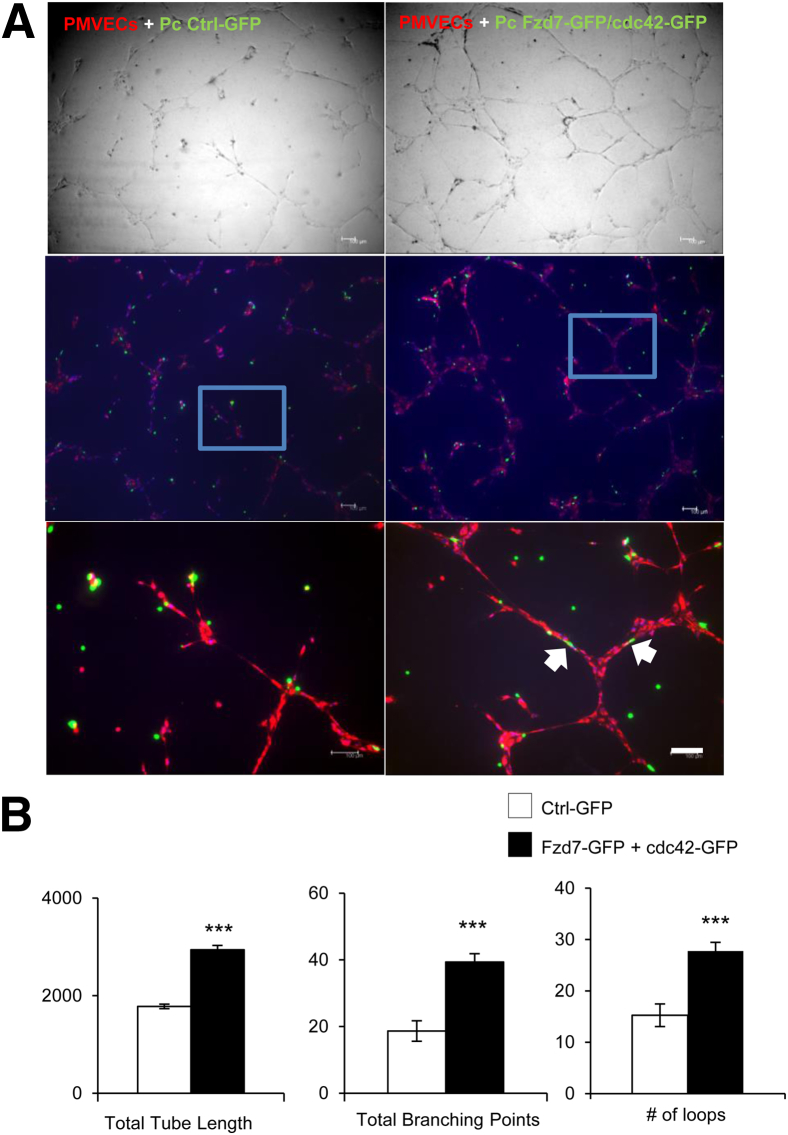
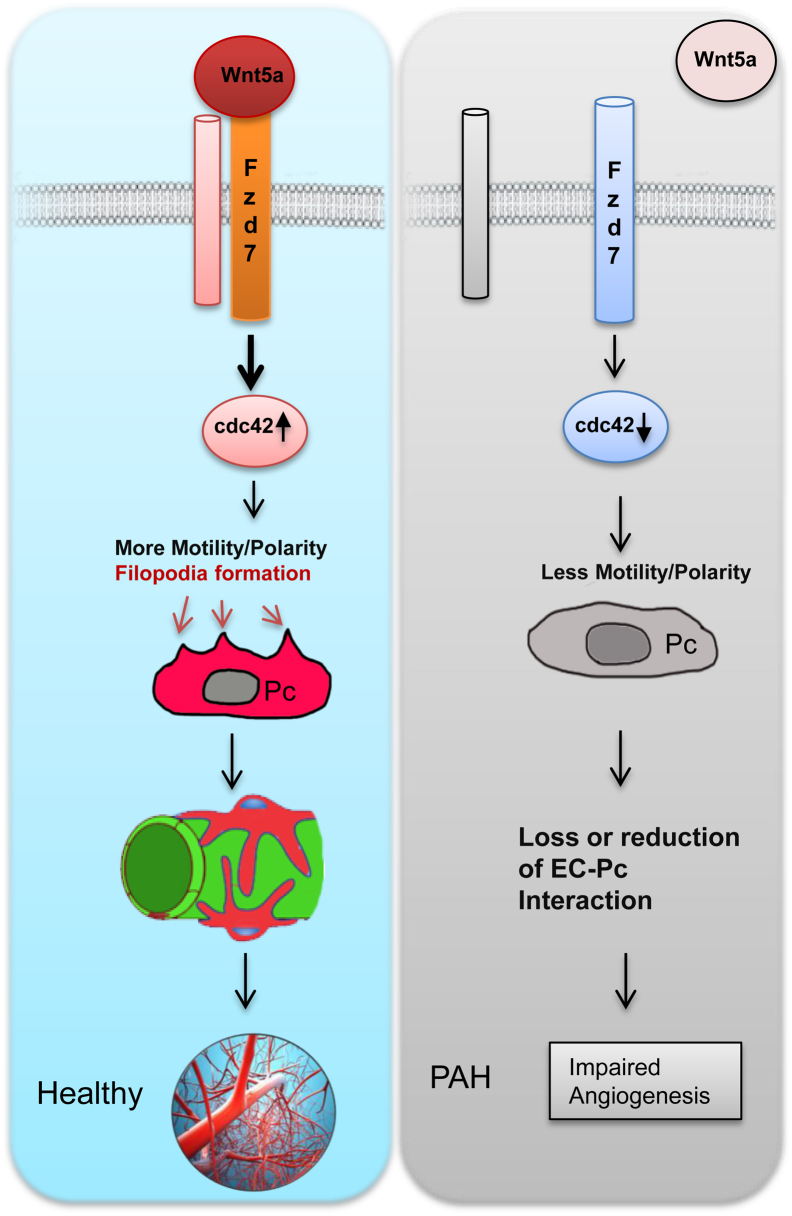
Pericytes are perivascular cells localized to capillaries that promote vessel maturation, and their absence can contribute to vessel loss. Whether impaired endothelial-pericyte interaction contributes to small vessel loss in pulmonary arterial hypertension (PAH) is unclear. Using 3G5-specific, immunoglobulin G-coated magnetic beads, we isolated pericytes from the lungs of healthy subjects and PAH patients, followed by lineage validation. PAH pericytes seeded with healthy pulmonary microvascular endothelial cells failed to associate with endothelial tubes, resulting in smaller vascular networks compared to those with healthy pericytes. After the demonstration of abnormal polarization toward endothelium via live-imaging and wound-healing studies, we screened PAH pericytes for abnormalities in the Wnt/planar cell polarity (PCP) pathway, which has been shown to regulate cell motility and polarity in the pulmonary vasculature. PAH pericytes had reduced expression of frizzled 7 (Fzd7) and cdc42, genes crucial for Wnt/PCP activation. With simultaneous knockdown of Fzd7 and cdc42 in healthy pericytes in vitro and in a murine model of angiogenesis, motility and polarization toward pulmonary microvascular endothelial cells were reduced, whereas with restoration of both genes in PAH pericytes, endothelial-pericyte association was improved, with larger vascular networks. These studies suggest that the motility and polarity of pericytes during pulmonary angiogenesis are regulated by Wnt/PCP activation, which can be targeted to prevent vessel loss in PAH.
Copyright © 2015 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Ribatti D., Nico B., Crivellato E. The role of pericytes in angiogenesis. Int J Dev Biol. 2011;55:261–268. - PubMed
-
- Yamagishi S., Imaizumi T. Pericyte biology and diseases. Int J Tissue React. 2005;27:125–135. - PubMed
-
- Diaz-Flores L., Gutierrez R., Madrid J.F., Varela H., Valladares F., Acosta E., Martin-Vasallo P., Diaz-Flores L., Jr. Pericytes. Morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche. Histol Histopathol. 2009;24:909–969. - PubMed
-
- Armulik A., Abramsson A., Betsholtz C. Endothelial/pericyte interactions. Circ Res. 2005;97:512–523. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
