

Ethanol at low concentrations protects glomerular podocytes through alcohol dehydrogenase and 20-HETE
- PMID: 25447342
- PMCID: PMC4385495
- DOI: 10.1016/j.prostaglandins.2014.10.006
Ethanol at low concentrations protects glomerular podocytes through alcohol dehydrogenase and 20-HETE
Abstract
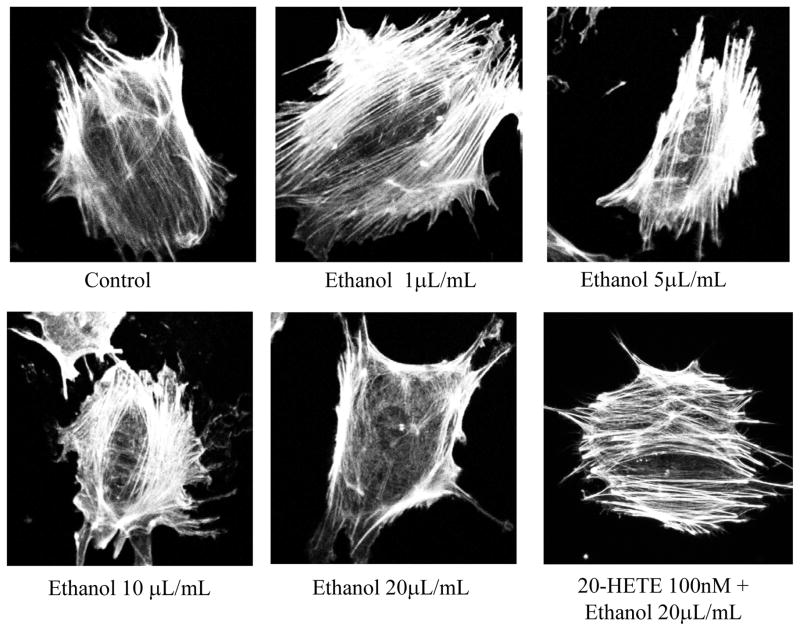
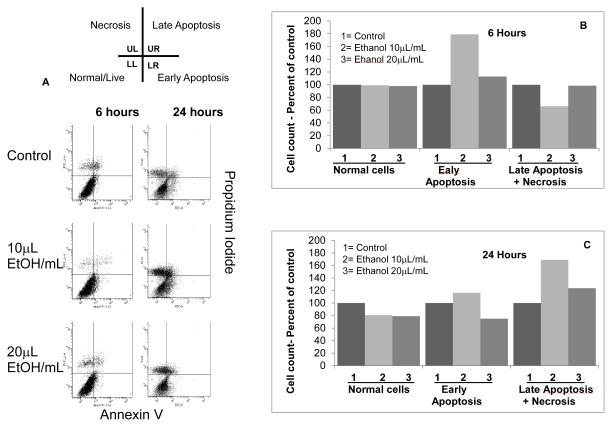
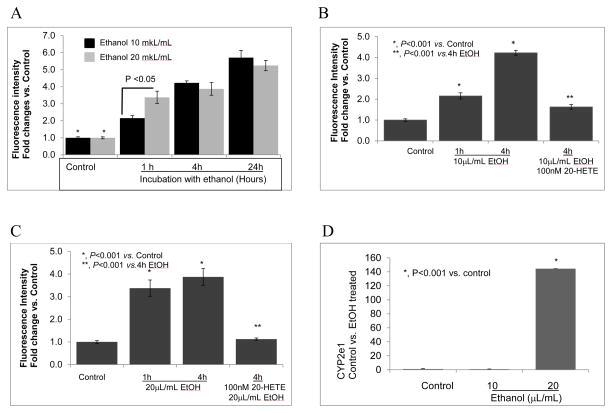
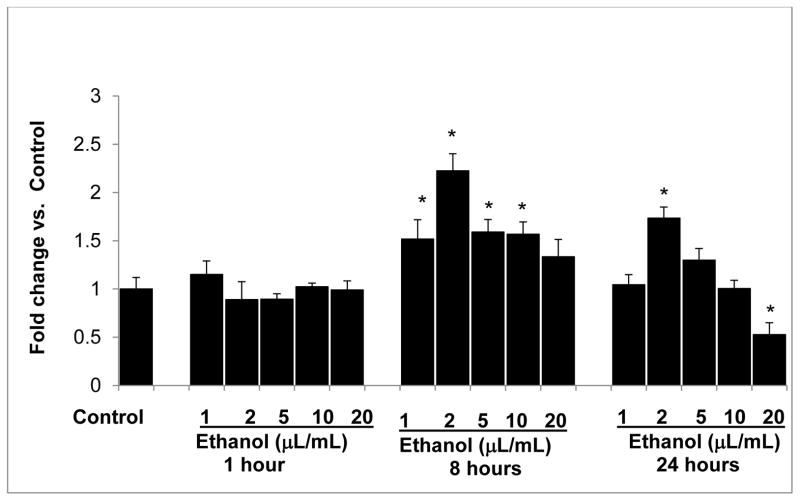
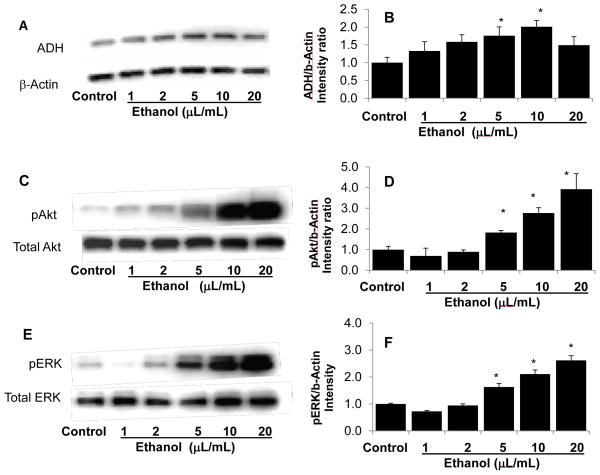
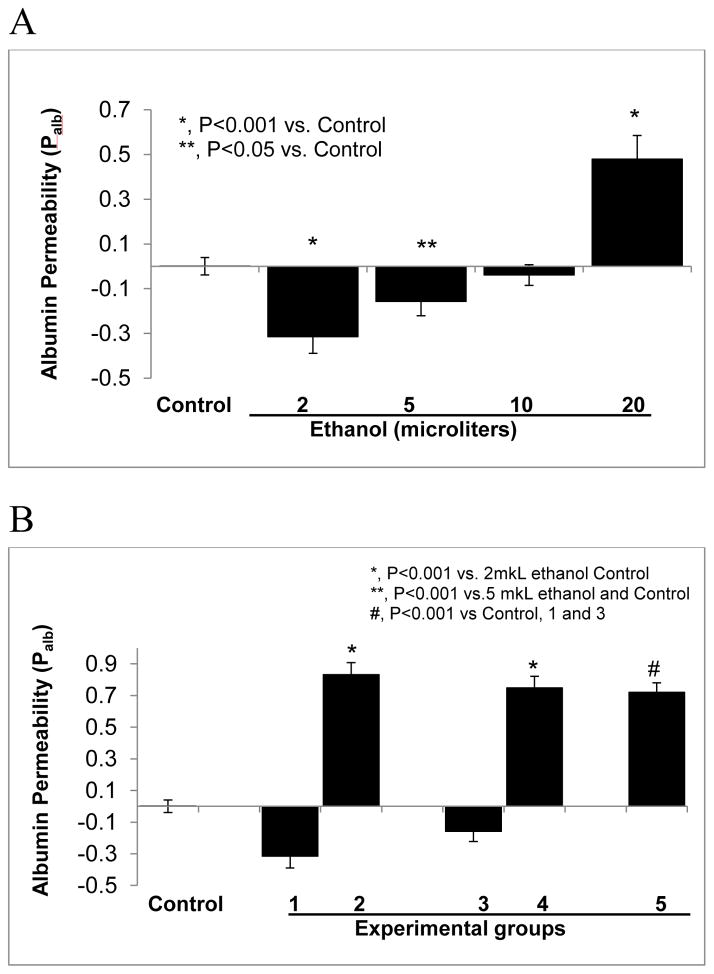
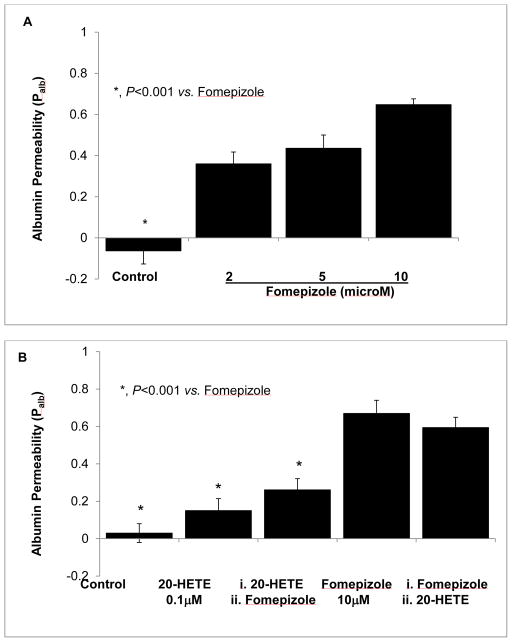
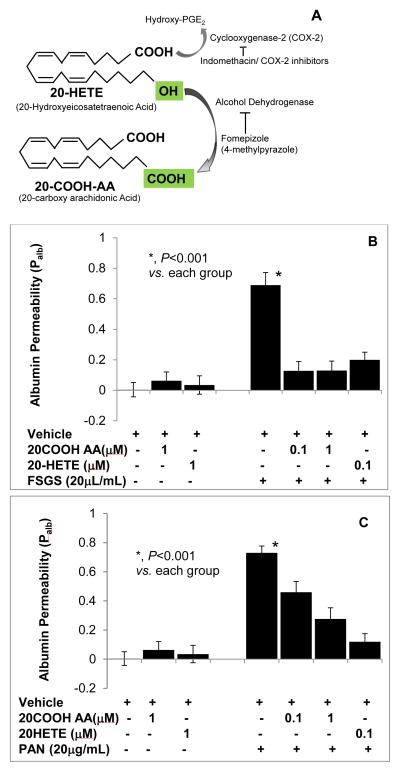
Clinical studies suggest cardiovascular and renal benefits of ingesting small amounts of ethanol. Effects of ethanol, role of alcohol dehydrogenase (ADH) or of 20-hydroxyeicosatetraenoic acid (20-HETE) in podocytes of the glomerular filtration barrier have not been reported. We found that mouse podocytes at baseline generate 20-HETE and express ADH but not CYP2e1. Ethanol at high concentrations altered the actin cytoskeleton, induced CYP2e1, increased superoxide production and inhibited ADH gene expression. Ethanol at low concentrations upregulated the expression of ADH and CYP4a12a. 20-HETE, an arachidonic acid metabolite generated by CYP4a12a, blocked the ethanol-induced cytoskeletal derangement and superoxide generation. Ethanol at high concentration or ADH inhibitor increased glomerular albumin permeability in vitro. 20-HETE and its metabolite produced by ADH activity, 20-carboxy-arachidonic acid, protected the glomerular permeability barrier against an ADH inhibitor, puromycin or FSGS permeability factor. We conclude that ADH activity is required for glomerular function, 20-HETE is a physiological substrate of ADH in podocytes and that podocytes are useful biosensors to understand glomeruloprotective effects of ethanol.
Keywords: 20-Carboxy-arachidonic acid; 20-Hydroxyeicosatetraenoic Acid; Alcohol dehydrogenase; Chronic kidney disease; Ethanol; Glomerular filtration barrier; Oxidative stress; Podocytes; Proteinuria.
Published by Elsevier Inc.
Conflict of interest statement
None of the authors have any conflict of interest to declare regarding the contents of the paper.
Figures
References
-
- Lenfant C, Chobanian AV, Jones DW, Roccella EJ. Seventh report of the Joint National Committee on the Prevention, Detection, Evaluation, and Treatment of High Blood Pressure (JNC 7): resetting the hypertension sails. Hypertension. 2003;41:1178–9. - PubMed
-
- Thun MJ, Peto R, Lopez AD, Monaco JH, Henley SJ, Heath CW, Jr, et al. Alcohol consumption and mortality among middle-aged and elderly U.S. adults. N Engl J Med. 1997;337:1705–14. - PubMed
-
- Di Castelnuovo A, Costanzo S, Bagnardi V, Donati MB, Iacoviello L, de Gaetano G. Alcohol dosing and total mortality in men and women: an updated meta-analysis of 34 prospective studies. Arch Intern Med. 2006;166:2437–45. - PubMed
-
- Schaeffner E, Ritz E. Alcohol and kidney damage: a Janus-faced relationship. Kidney Int. 2012;81:816–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
