

Alcohol disrupts levels and function of the cystic fibrosis transmembrane conductance regulator to promote development of pancreatitis
- PMID: 25447846
- PMCID: PMC4353632
- DOI: 10.1053/j.gastro.2014.11.002
Alcohol disrupts levels and function of the cystic fibrosis transmembrane conductance regulator to promote development of pancreatitis
Abstract
Background & aims: Excessive consumption of ethanol is one of the most common causes of acute and chronic pancreatitis. Alterations to the gene encoding the cystic fibrosis transmembrane conductance regulator (CFTR) also cause pancreatitis. However, little is known about the role of CFTR in the pathogenesis of alcohol-induced pancreatitis.
Methods: We measured CFTR activity based on chloride concentrations in sweat from patients with cystic fibrosis, patients admitted to the emergency department because of excessive alcohol consumption, and healthy volunteers. We measured CFTR levels and localization in pancreatic tissues and in patients with acute or chronic pancreatitis induced by alcohol. We studied the effects of ethanol, fatty acids, and fatty acid ethyl esters on secretion of pancreatic fluid and HCO3(-), levels and function of CFTR, and exchange of Cl(-) for HCO3(-) in pancreatic cell lines as well as in tissues from guinea pigs and CFTR knockout mice after administration of alcohol.
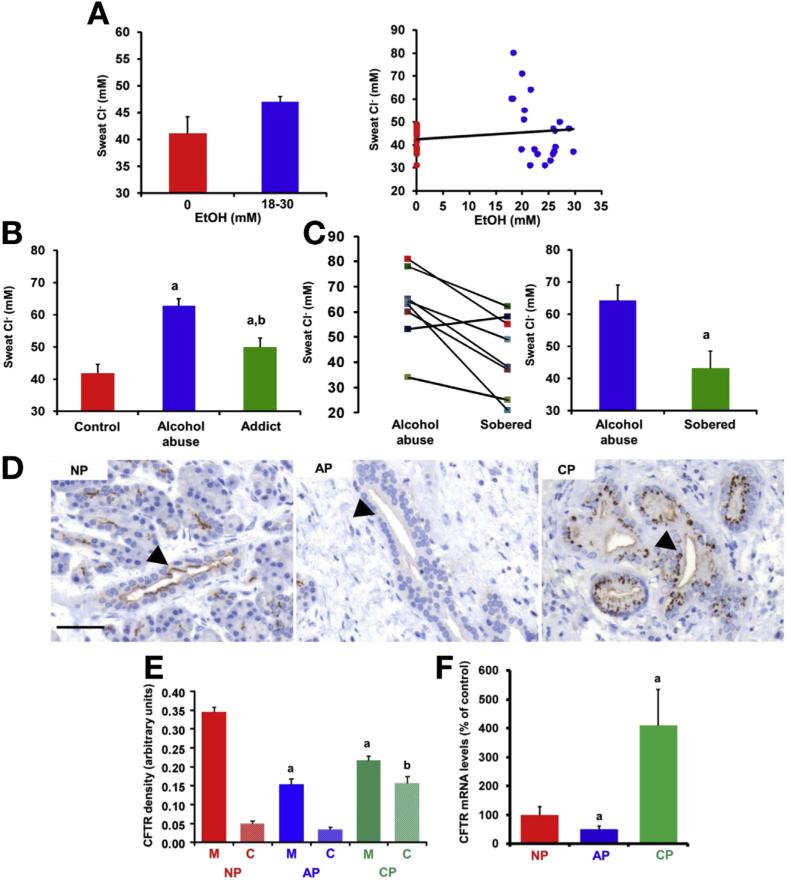
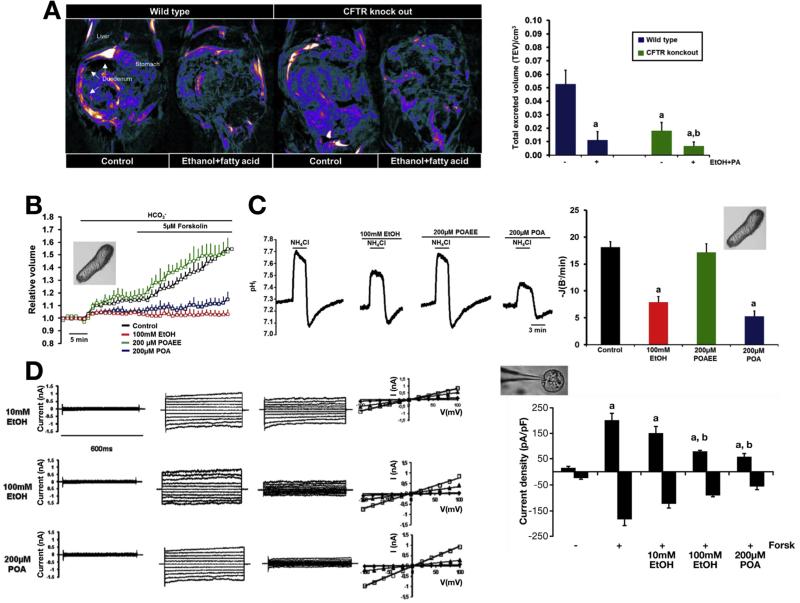
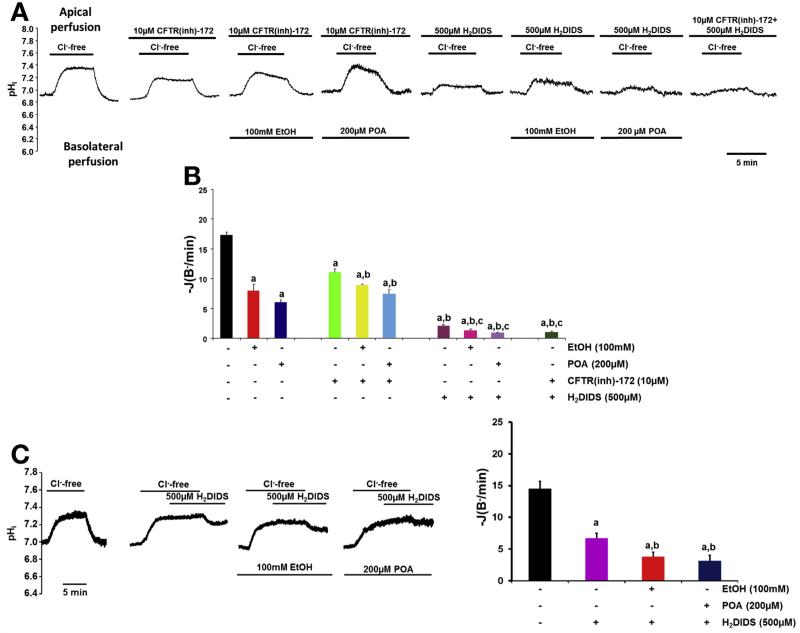
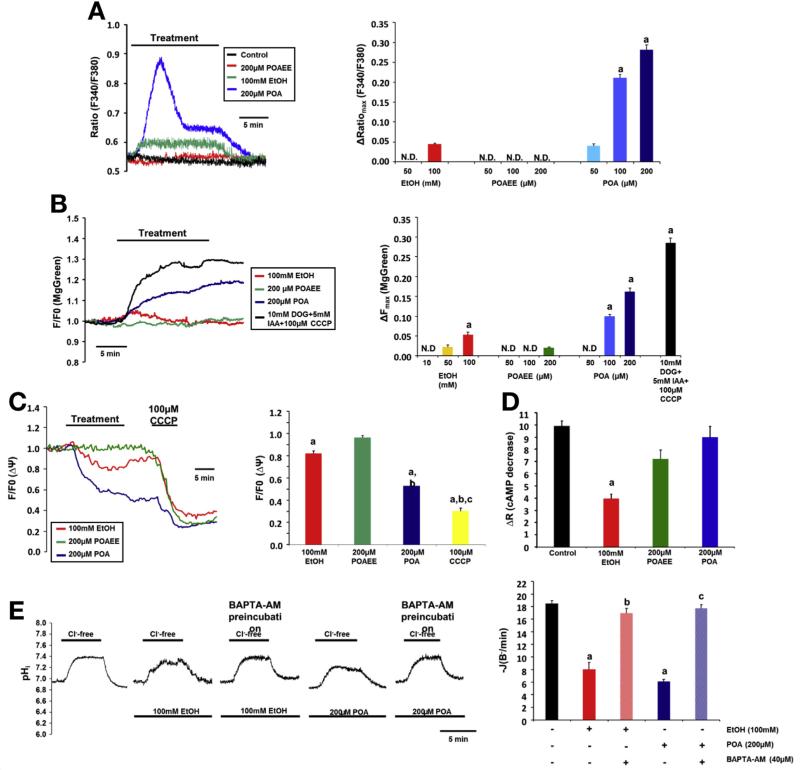
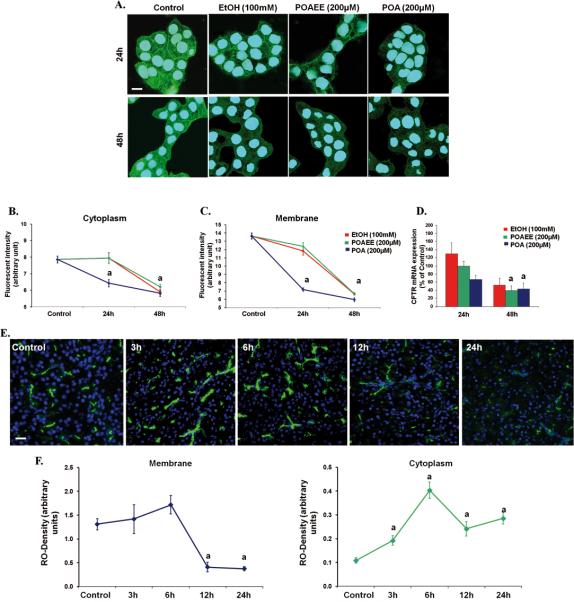
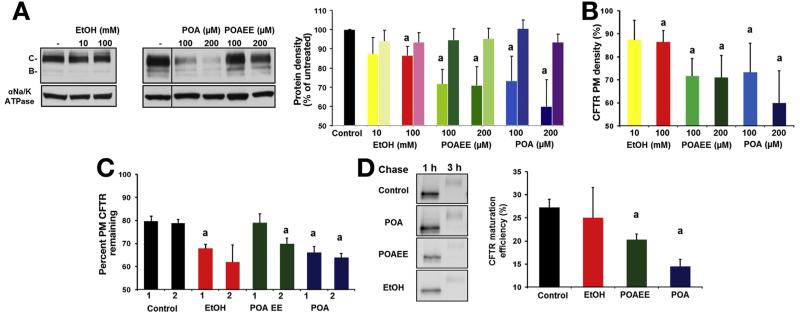
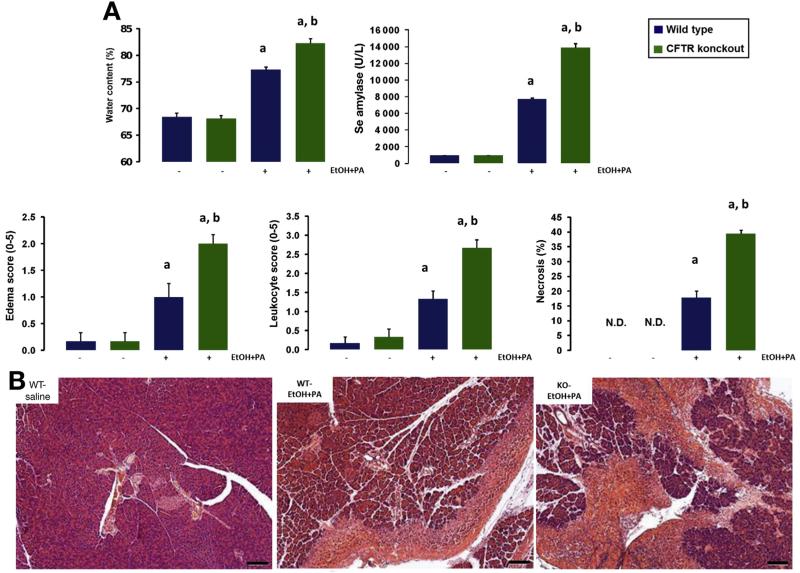
Results: Chloride concentrations increased in sweat samples from patients who acutely abused alcohol but not in samples from healthy volunteers, indicating that alcohol affects CFTR function. Pancreatic tissues from patients with acute or chronic pancreatitis had lower levels of CFTR than tissues from healthy volunteers. Alcohol and fatty acids inhibited secretion of fluid and HCO3(-), as well as CFTR activity, in pancreatic ductal epithelial cells. These effects were mediated by sustained increases in concentrations of intracellular calcium and adenosine 3',5'-cyclic monophosphate, depletion of adenosine triphosphate, and depolarization of mitochondrial membranes. In pancreatic cell lines and pancreatic tissues of mice and guinea pigs, administration of ethanol reduced expression of CFTR messenger RNA, reduced the stability of CFTR at the cell surface, and disrupted folding of CFTR at the endoplasmic reticulum. CFTR knockout mice given ethanol or fatty acids developed more severe pancreatitis than mice not given ethanol or fatty acids.
Conclusions: Based on studies of human, mouse, and guinea pig pancreata, alcohol disrupts expression and localization of the CFTR. This appears to contribute to development of pancreatitis. Strategies to increase CFTR levels or function might be used to treat alcohol-associated pancreatitis.
Keywords: Alcoholism; Cl(−) Channel; Duct; Exocrine Pancreas.
Copyright © 2015 AGA Institute. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Pancreatitis: mislocalization of CFTR by alcohol promotes pancreatitis.Nat Rev Gastroenterol Hepatol. 2015 Jan;12(1):2. doi: 10.1038/nrgastro.2014.204. Epub 2014 Nov 25. Nat Rev Gastroenterol Hepatol. 2015. PMID: 25421582 No abstract available.
References
-
- Nagar AB, Gorelick FS. Acute pancreatitis. Curr Opin Gastroenterol. 2002;18:552–557. - PubMed
-
- Braganza JM, Lee SH, McCloy RF, et al. Chronic pancreatitis. Lancet. 2011;377:1184–1197. - PubMed
-
- Petersen OH, Tepikin AV, Gerasimenko JV, et al. Fatty acids, alcohol and fatty acid ethyl esters: toxic Ca2+ signal generation and pancreatitis. Cell Calcium. 2009;45:634–642. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
