

Luteolin inhibits Cr(VI)-induced malignant cell transformation of human lung epithelial cells by targeting ROS mediated multiple cell signaling pathways
- PMID: 25448439
- PMCID: PMC4433031
- DOI: 10.1016/j.taap.2014.10.008
Luteolin inhibits Cr(VI)-induced malignant cell transformation of human lung epithelial cells by targeting ROS mediated multiple cell signaling pathways
Abstract
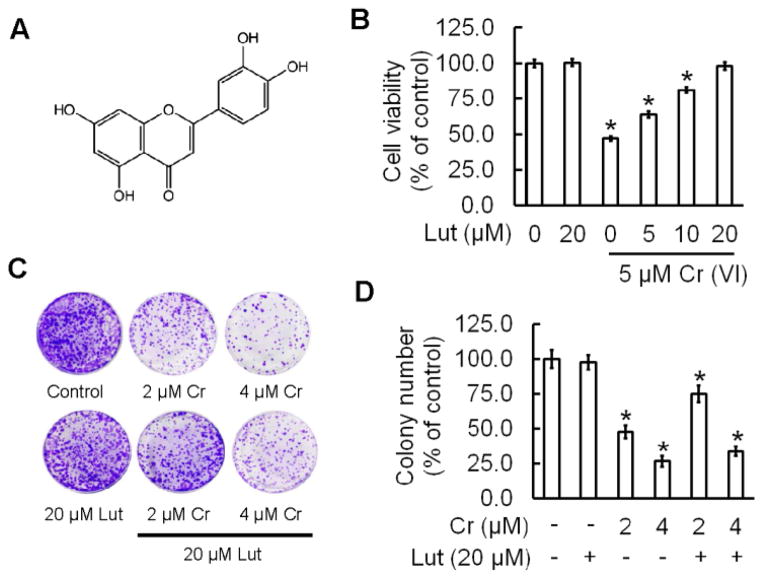
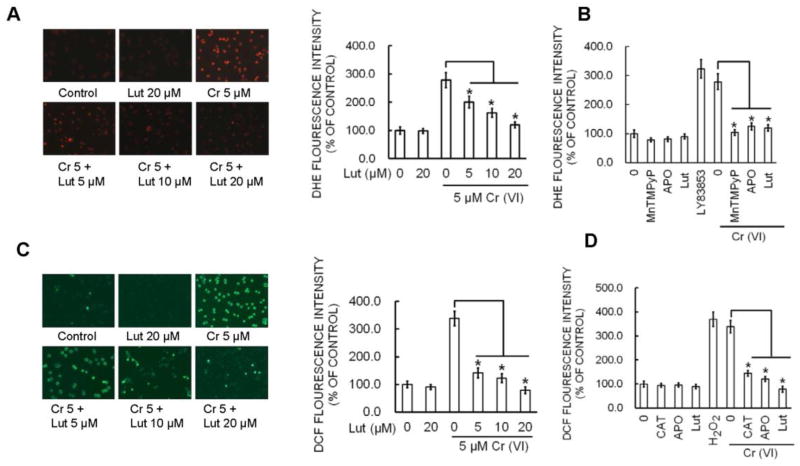
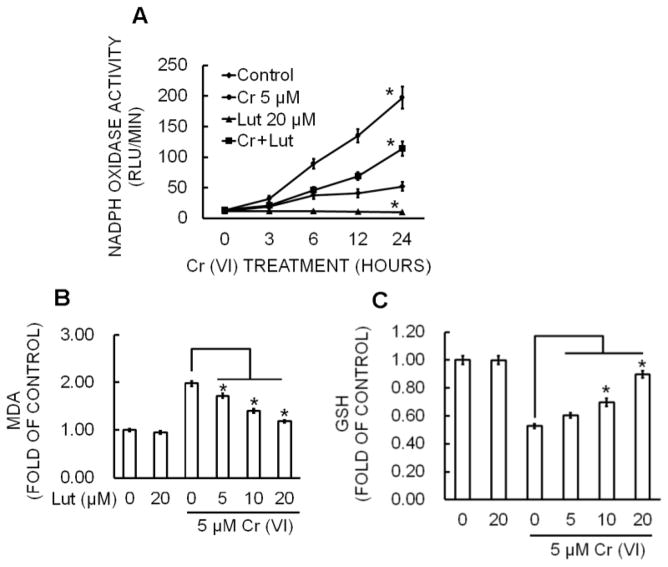
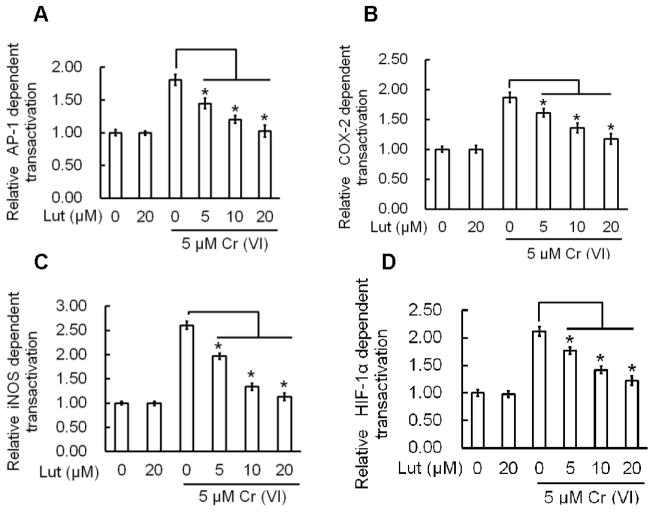
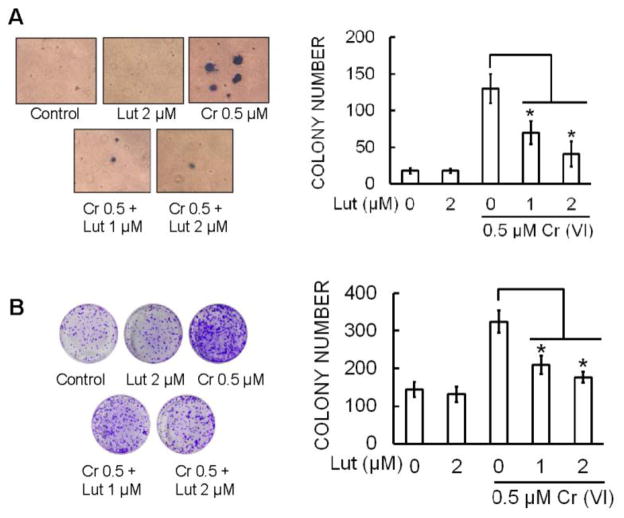
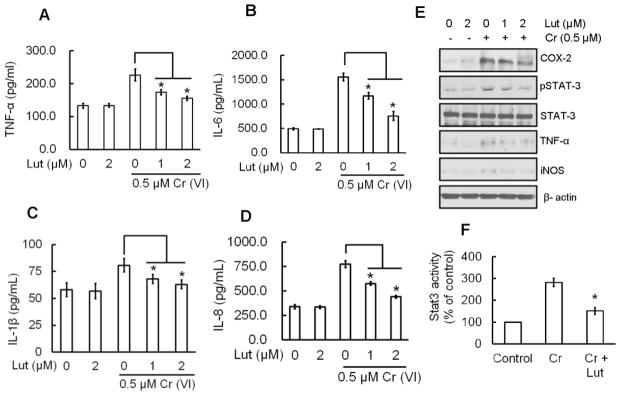
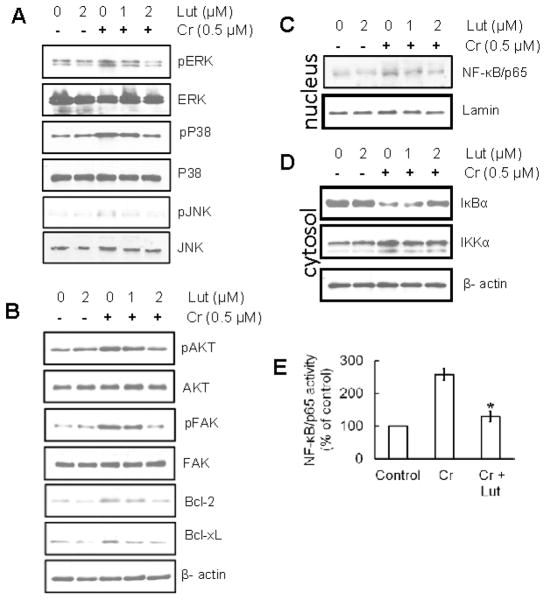
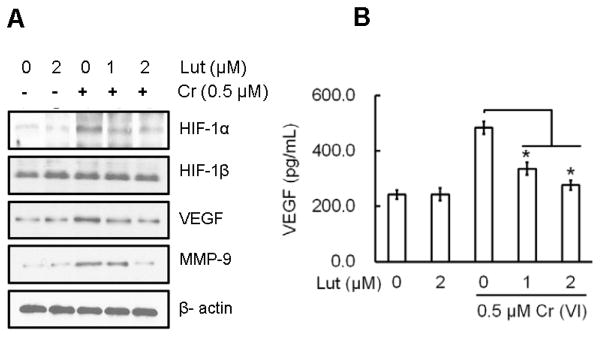
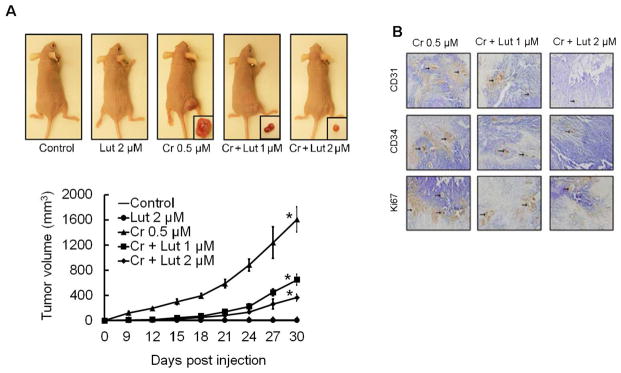
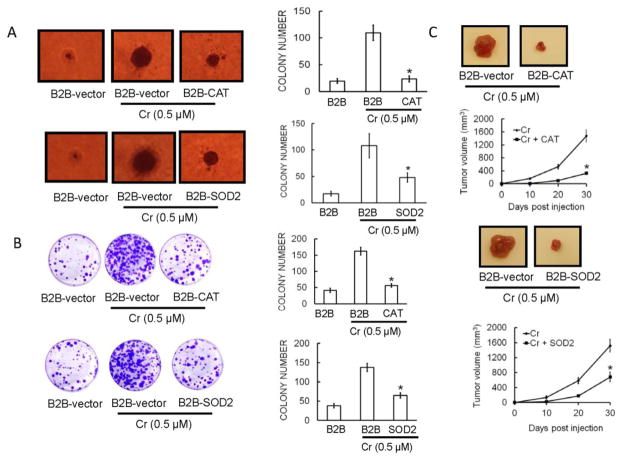
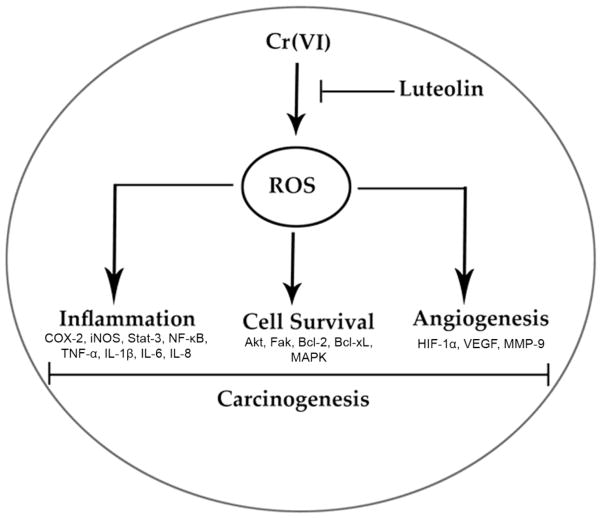
Hexavalent chromium [Cr(VI)] is a well-known human carcinogen associated with the incidence of lung cancer. Inhibition of metal induced carcinogenesis by a dietary antioxidant is a novel approach. Luteolin, a natural dietary flavonoid found in fruits and vegetables, possesses potent antioxidant and anti-inflammatory activity. We found that short term exposure of human bronchial epithelial cells (BEAS-2B) to Cr(VI) (5μM) showed a drastic increase in ROS generation, NADPH oxidase (NOX) activation, lipid peroxidation, and glutathione depletion, which were significantly inhibited by the treatment with luteolin in a dose dependent manner. Treatment with luteolin decreased AP-1, HIF-1α, COX-2, and iNOS promoter activity induced by Cr(VI) in BEAS-2B cells. In addition, luteolin protected BEAS-2B cells from malignant transformation induced by chronic Cr(VI) exposure. Moreover, luteolin also inhibited the production of pro-inflammatory cytokines (IL-1β, IL-6, IL-8, TNF-α) and VEGF in chronic Cr(VI) exposed BEAS-2B cells. Western blot analysis showed that luteolin inhibited multiple gene products linked to survival (Akt, Fak, Bcl-2, Bcl-xL), inflammation (MAPK, NF-κB, COX-2, STAT-3, iNOS, TNF-α) and angiogenesis (HIF-1α, VEGF, MMP-9) in chronic Cr(VI) exposed BEAS-2B cells. Nude mice injected with BEAS-2B cells chronically exposed to Cr(VI) in the presence of luteolin showed reduced tumor incidence compared to Cr(VI) alone treated group. Overexpression of catalase (CAT) or SOD2, eliminated Cr(VI)-induced malignant transformation. Overall, our results indicate that luteolin protects BEAS-2B cells from Cr(VI)-induced carcinogenesis by scavenging ROS and modulating multiple cell signaling mechanisms that are linked to ROS. Luteolin, therefore, serves as a potential chemopreventive agent against Cr(VI)-induced carcinogenesis.
Keywords: Angiogenesis; Carcinogenesis; Hexavalent chromium; Inflammation; Luteolin.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
Disclosure of Potential Conflicts of Interest: No potential conflicts of interest were disclosed.
Figures
Similar articles
-
Protection from Cr(VI)-induced malignant cell transformation and tumorigenesis of Cr(VI)-transformed cells by luteolin through Nrf2 signaling.Toxicol Appl Pharmacol. 2017 Sep 15;331:24-32. doi: 10.1016/j.taap.2017.04.016. Epub 2017 Apr 14. Toxicol Appl Pharmacol. 2017. PMID: 28416455 Free PMC article.
-
Hexavalent chromium induces malignant transformation of human lung bronchial epithelial cells via ROS-dependent activation of miR-21-PDCD4 signaling.Oncotarget. 2016 Aug 9;7(32):51193-51210. doi: 10.18632/oncotarget.9967. Oncotarget. 2016. PMID: 27323401 Free PMC article.
-
NADPH oxidase activation is required in reactive oxygen species generation and cell transformation induced by hexavalent chromium.Toxicol Sci. 2011 Oct;123(2):399-410. doi: 10.1093/toxsci/kfr180. Epub 2011 Jul 8. Toxicol Sci. 2011. PMID: 21742780 Free PMC article.
-
Assessment of the mode of action for hexavalent chromium-induced lung cancer following inhalation exposures.Toxicology. 2014 Nov 5;325:160-79. doi: 10.1016/j.tox.2014.08.009. Epub 2014 Aug 28. Toxicology. 2014. PMID: 25174529 Review.
-
Oxidative stress and chromium(VI) carcinogenesis.J Environ Pathol Toxicol Oncol. 2008;27(2):77-88. doi: 10.1615/jenvironpatholtoxicoloncol.v27.i2.10. J Environ Pathol Toxicol Oncol. 2008. PMID: 18540844 Review.
Cited by
-
Molecular and epigenetic mechanisms of Cr(VI)-induced carcinogenesis.Toxicol Appl Pharmacol. 2019 Aug 15;377:114636. doi: 10.1016/j.taap.2019.114636. Epub 2019 Jun 20. Toxicol Appl Pharmacol. 2019. PMID: 31228494 Free PMC article. Review.
-
Protection from Cr(VI)-induced malignant cell transformation and tumorigenesis of Cr(VI)-transformed cells by luteolin through Nrf2 signaling.Toxicol Appl Pharmacol. 2017 Sep 15;331:24-32. doi: 10.1016/j.taap.2017.04.016. Epub 2017 Apr 14. Toxicol Appl Pharmacol. 2017. PMID: 28416455 Free PMC article.
-
Quercetin potentializes the respective cytotoxic activity of gemcitabine or doxorubicin on 3D culture of AsPC-1 or HepG2 cells, through the inhibition of HIF-1α and MDR1.PLoS One. 2020 Oct 14;15(10):e0240676. doi: 10.1371/journal.pone.0240676. eCollection 2020. PLoS One. 2020. PMID: 33052979 Free PMC article.
-
A Newly Synthesized Flavone from Luteolin Escapes from COMT-Catalyzed Methylation and Inhibits Lipopolysaccharide-Induced Inflammation in RAW264.7 Macrophages via JNK, p38 and NF-κB Signaling Pathways.J Microbiol Biotechnol. 2022 Jan 28;32(1):15-26. doi: 10.4014/jmb.2104.04027. J Microbiol Biotechnol. 2022. PMID: 34099595 Free PMC article.
-
Hexavalent chromium induces malignant transformation of human lung bronchial epithelial cells via ROS-dependent activation of miR-21-PDCD4 signaling.Oncotarget. 2016 Aug 9;7(32):51193-51210. doi: 10.18632/oncotarget.9967. Oncotarget. 2016. PMID: 27323401 Free PMC article.
References
-
- Abreu P, Ferreira L, Alpoim M, Urbano A. Impact of hexavalent chromium on mammalian cell bioenergetics: phenotypic changes, molecular basis and potential relevance to chromate-induced lung cancer. BioMetals. 2014;27:409–443. - PubMed
-
- Ahmad MK, Syma S, Mahmood R. Cr (VI) induces lipid peroxidation, protein oxidation and alters the activities of antioxidant enzymes in human erythrocytes. Biological trace element research. 2011;144:426–435. - PubMed
-
- Anand SS. Protective effect of vitamin B6 in chromium-induced oxidative stress in liver. Journal of Applied Toxicology. 2005;25:440–443. - PubMed
-
- Azad N, Rojanasakul Y, Vallyathan V. Inflammation and lung cancer: roles of reactive oxygen/nitrogen species. Journal of Toxicology and Environmental Health, Part B. 2008;11:1–15. - PubMed
-
- Ballif BA, Blenis J. Molecular mechanisms mediating mammalian mitogen-activated protein kinase (MAPK) kinase (MEK)-MAPK cell survival signals. Cell growth and differentiation. 2001;12:397–408. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
