

AMPK--sensing energy while talking to other signaling pathways
- PMID: 25448702
- PMCID: PMC5693325
- DOI: 10.1016/j.cmet.2014.09.013
AMPK--sensing energy while talking to other signaling pathways
Abstract
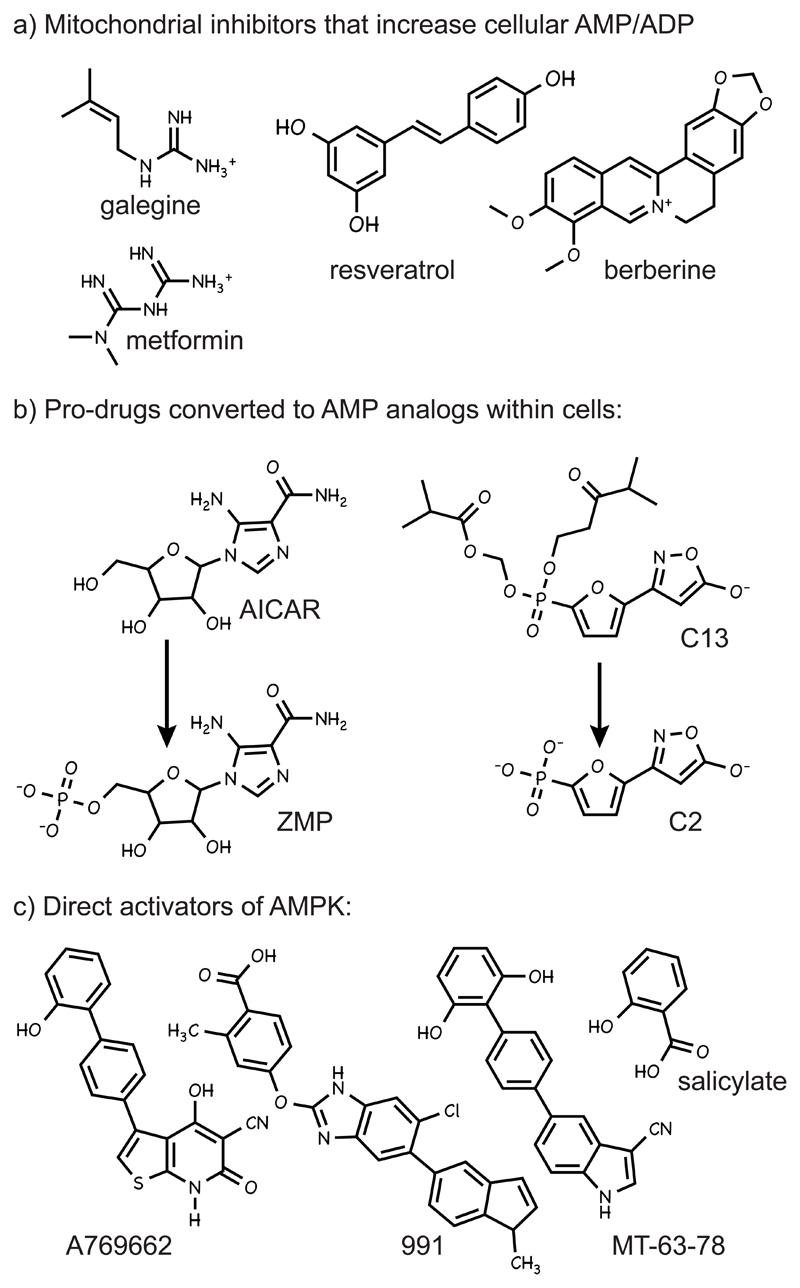
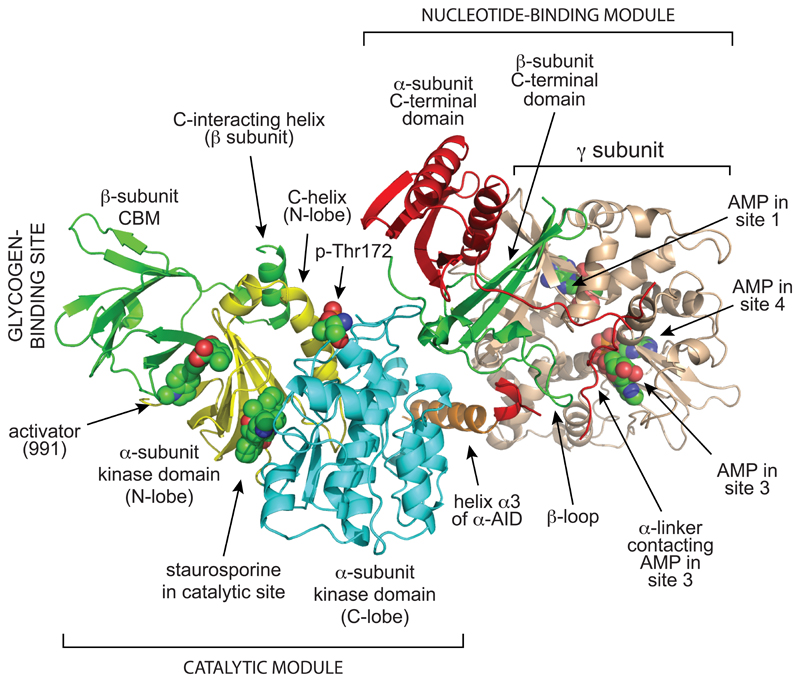
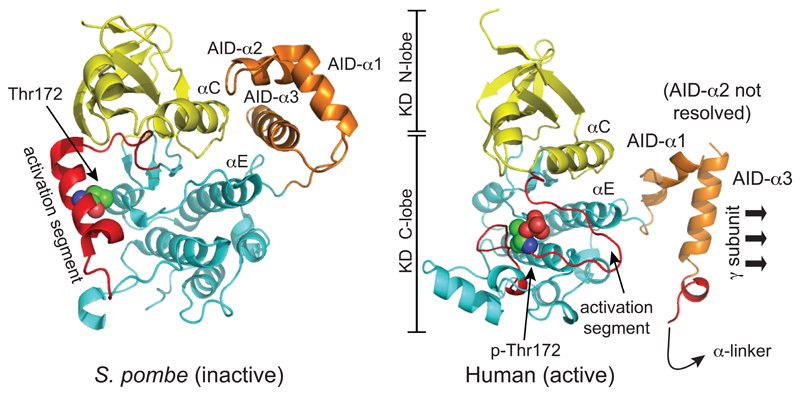
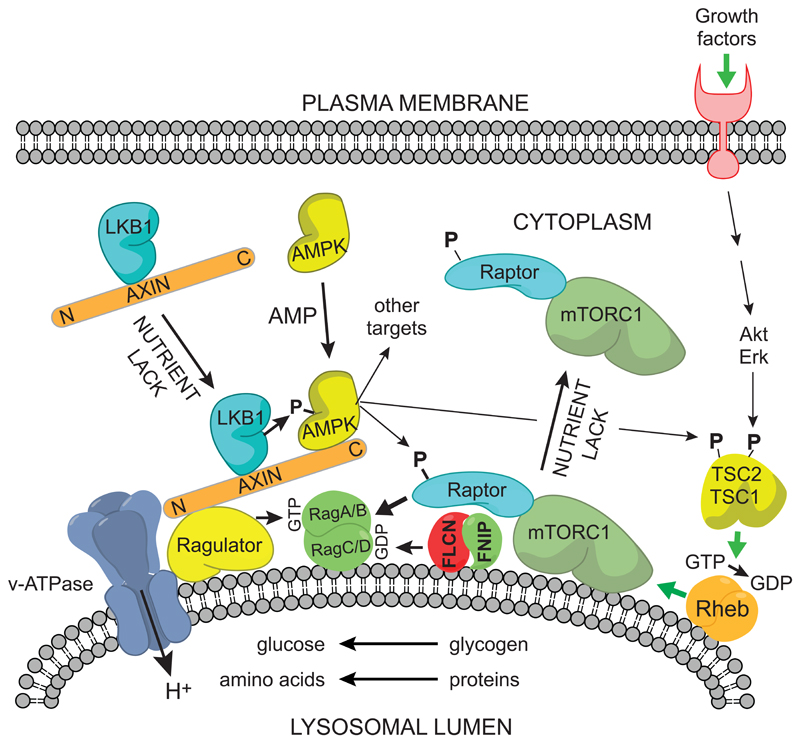
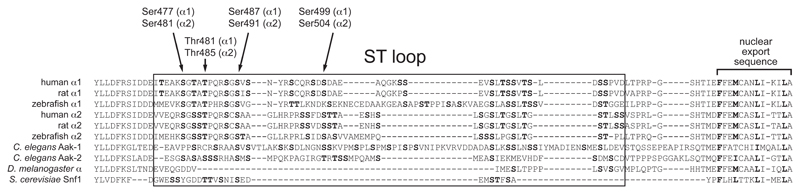
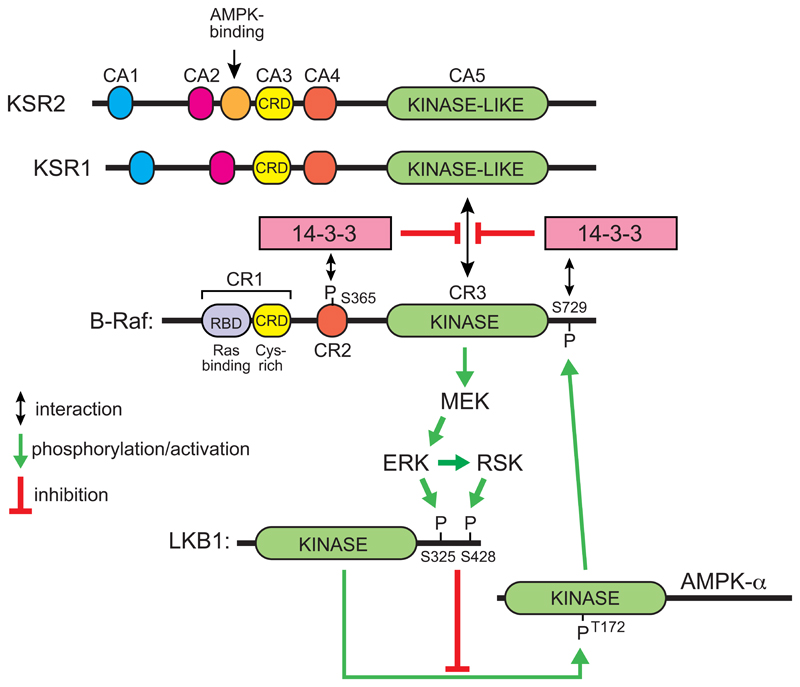
The AMP-activated protein kinase (AMPK) is a sensor of cellular energy and nutrient status, expressed almost universally in eukaryotes as heterotrimeric complexes comprising catalytic (α) and regulatory (β and γ) subunits. Along with the mechanistic target of rapamycin complex-1 (mTORC1), AMPK may have been one of the earliest signaling pathways to have arisen during eukaryotic evolution. Recent crystal structures have provided insights into the mechanisms by which AMPK is regulated by phosphorylation and allosteric activators. Another recent development has been the realization that activation of AMPK by the upstream kinase LKB1 may primarily occur not in the cytoplasm, but at the surface of the lysosome, where AMPK and mTORC1 are regulated in a reciprocal manner by the availability of nutrients. It is also becoming clear that there is a substantial amount of crosstalk between the AMPK pathway and other signaling pathways that promote cell growth and proliferation, and this will be discussed.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Amodeo GA, Rudolph MJ, Tong L. Crystal structure of the heterotrimer core of Saccharomyces cerevisiae AMPK homologue SNF1. Nature. 2007;449:492–495. - PubMed
-
- Baba M, Hong SB, Sharma N, Warren MB, Nickerson ML, Iwamatsu A, Esposito D, Gillette WK, Hopkins RF, 3rd, Hartley JL, et al. Folliculin encoded by the BHD gene interacts with a binding protein, FNIP1, and AMPK, and is involved in AMPK and mTOR signaling. Proc Natl Acad Sci U S A. 2006;103:15552–15557. - PMC - PubMed
-
- Bateman A. The structure of a domain common to archaebacteria and the homocystinuria disease protein. Trends Biochem Sci. 1997;22:12–13. - PubMed
-
- Bultot L, Guigas B, Von Wilamowitz-Moellendorff A, Maisin L, Vertommen D, Hussain N, Beullens M, Guinovart JJ, Foretz M, Viollet B, et al. AMP-activated protein kinase phosphorylates and inactivates liver glycogen synthase. Biochem J. 2012;443:193–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
