

Liver-specific deletion of augmenter of liver regeneration accelerates development of steatohepatitis and hepatocellular carcinoma in mice
- PMID: 25448926
- PMCID: PMC4802363
- DOI: 10.1053/j.gastro.2014.10.008
Liver-specific deletion of augmenter of liver regeneration accelerates development of steatohepatitis and hepatocellular carcinoma in mice
Abstract
Background & aims: Augmenter of liver regeneration (ALR, encoded by GFER) is a widely distributed pleiotropic protein originally identified as a hepatic growth factor. However, little is known about its roles in hepatic physiology and pathology. We created mice with liver-specific deletion of ALR to study its function.
Methods: We developed mice with liver-specific deletion of ALR (ALR-L-KO) using the albumin-Cre/LoxP system. Liver tissues were collected from ALR-L-KO mice and ALR(floxed/floxed) mice (controls) and analyzed by histology, reverse-transcription polymerase chain reaction, immunohistochemistry, electron microscopy, and techniques to measure fibrosis and lipids. Liver tissues from patients with and without advanced liver disease were determined by immunoblot analysis.
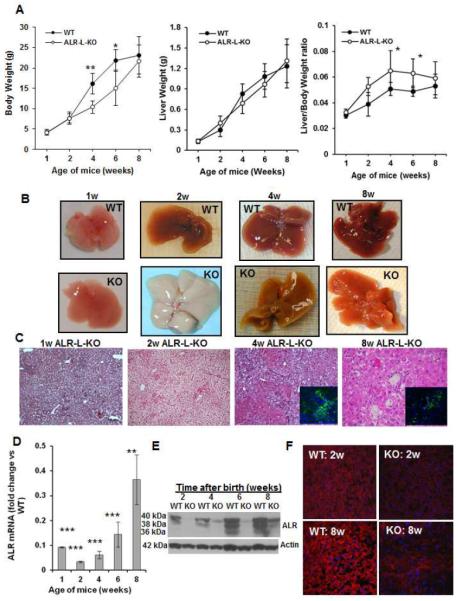
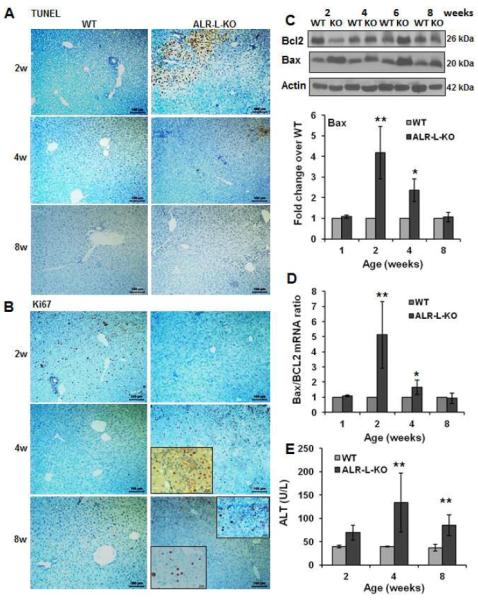
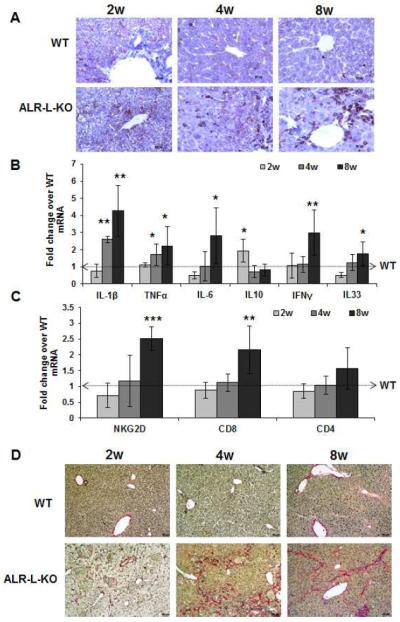
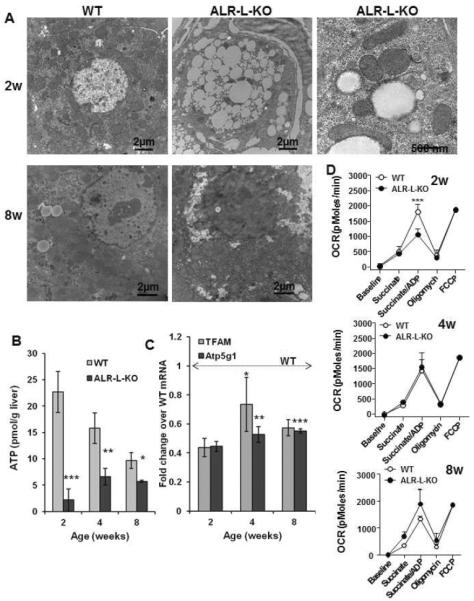
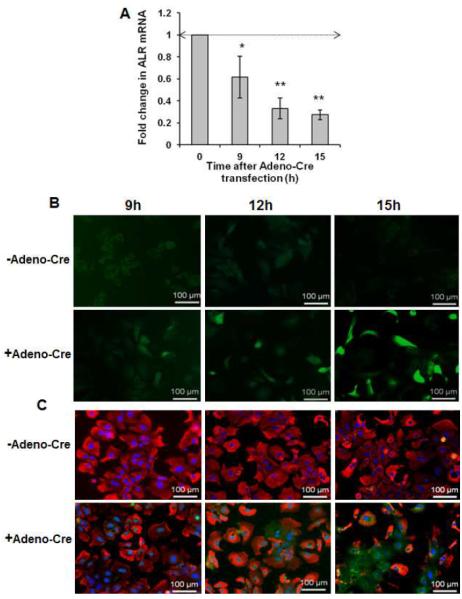
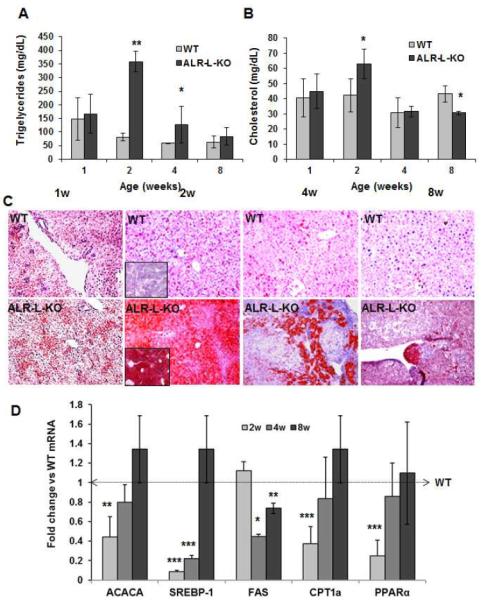
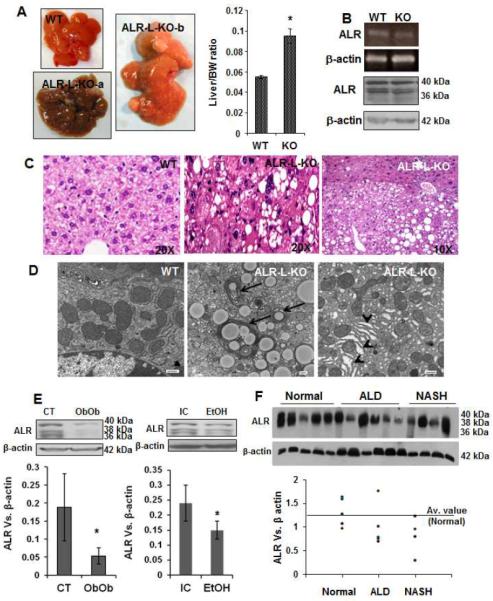
Results: Two weeks after birth, livers of ALR-L-KO mice contained low levels of ALR and adenosine triphosphate (ATP); they had reduced mitochondrial respiratory function and increased oxidative stress, compared with livers from control mice, and had excessive steatosis, and hepatocyte apoptosis. Levels of carbamyl-palmitoyl transferase 1a and ATP synthase subunit ATP5G1 were reduced in livers of ALR-L-KO mice, indicating defects in mitochondrial fatty acid transport and ATP synthesis. Electron microscopy showed mitochondrial swelling with abnormalities in shapes and numbers of cristae. From weeks 2-4 after birth, levels of steatosis and apoptosis decreased in ALR-L-KO mice, and numbers of ALR-expressing cells increased, along with ATP levels. However, at weeks 4-8 after birth, livers became inflamed, with hepatocellular necrosis, ductular proliferation, and fibrosis; hepatocellular carcinoma developed by 1 year after birth in nearly 60% of the mice. Hepatic levels of ALR were also low in ob/ob mice and alcohol-fed mice with liver steatosis, compared with controls. Levels of ALR were lower in liver tissues from patients with advanced alcoholic liver disease and nonalcoholic steatohepatitis than in control liver tissues.
Conclusions: We developed mice with liver-specific deletion of ALR, and showed that it is required for mitochondrial function and lipid homeostasis in the liver. ALR-L-KO mice provide a useful model for investigating the pathogenesis of steatohepatitis and its complications.
Keywords: ALD; Augmenter of Liver Regeneration; Mouse Model; NASH.
Copyright © 2015 AGA Institute. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Augmenter of liver regeneration links mitochondrial function to steatohepatitis and hepatocellular carcinoma.Gastroenterology. 2015 Feb;148(2):285-8. doi: 10.1053/j.gastro.2014.12.013. Epub 2014 Dec 18. Gastroenterology. 2015. PMID: 25529802 No abstract available.
References
-
- Francavilla A, Hagiya M, Porter KA, et al. Augmenter of liver regeneration: Its place in the universe of hepatic growth factors. Hepatology. 1994;20:747–757. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
