

Efficient backsplicing produces translatable circular mRNAs
- PMID: 25449546
- PMCID: PMC4338345
- DOI: 10.1261/rna.048272.114
Efficient backsplicing produces translatable circular mRNAs
Abstract
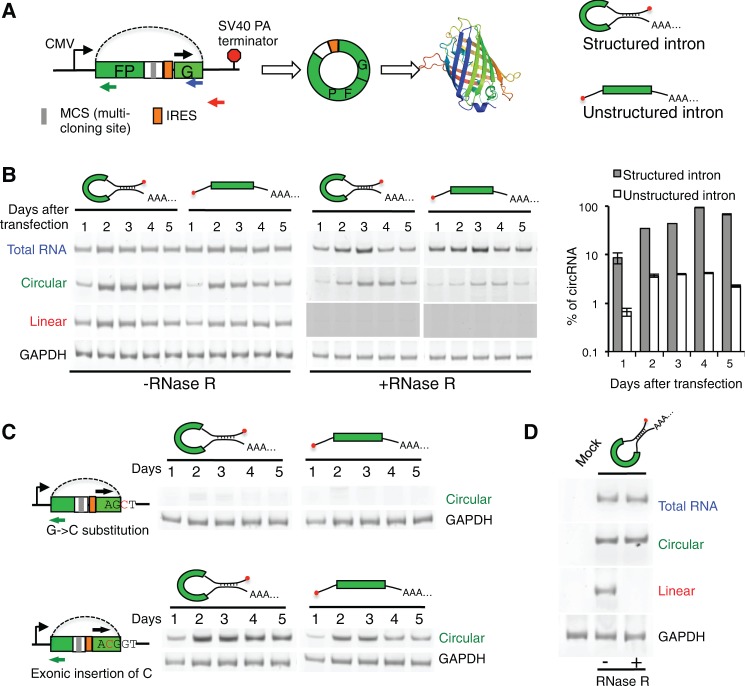
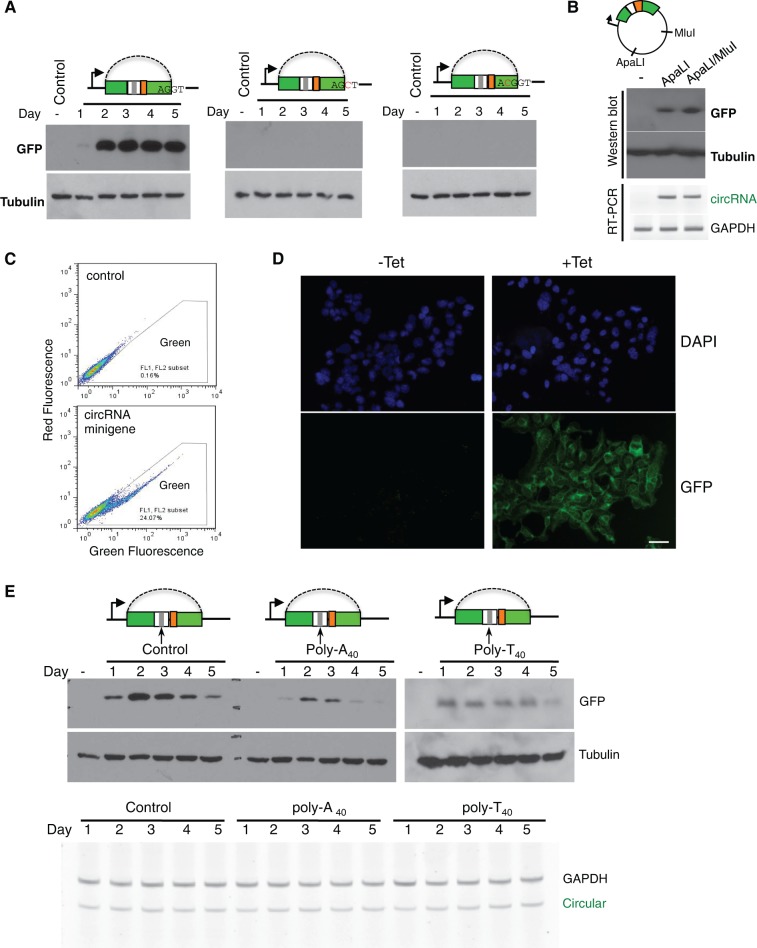
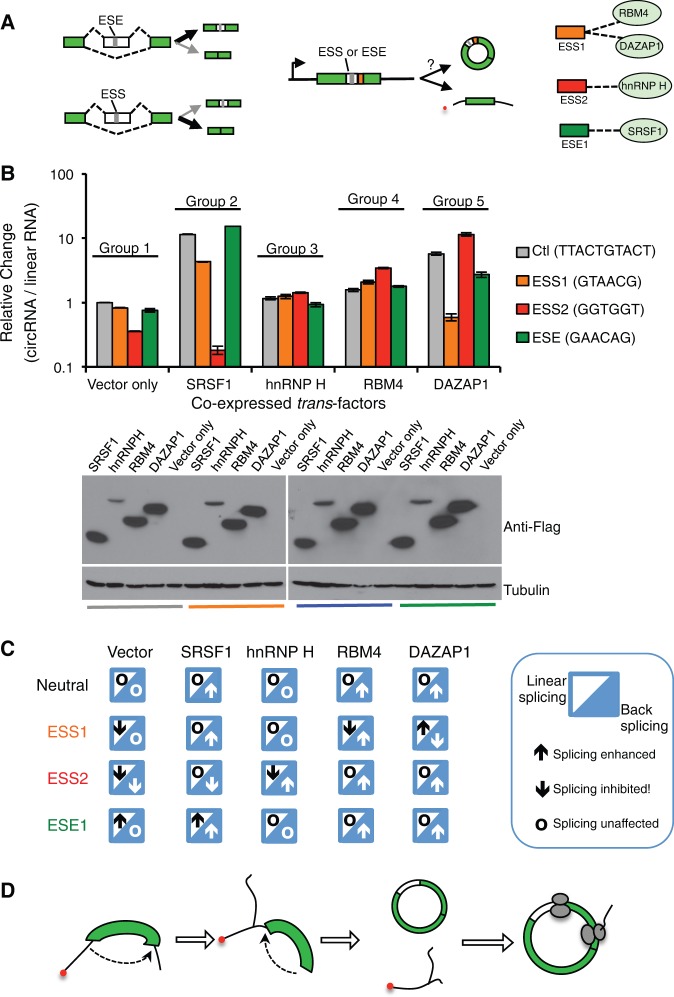
While the human transcriptome contains a large number of circular RNAs (circRNAs), the functions of most circRNAs remain unclear. Sequence annotation suggests that most circRNAs are generated from splicing in reversed orders across exons. However, the mechanisms of this backsplicing are largely unknown. Here we constructed a single exon minigene containing split GFP, and found that the pre-mRNA indeed produces circRNA through efficient backsplicing in human and Drosophila cells. The backsplicing is enhanced by complementary introns that form double-stranded RNA structure to bring splice sites in proximity, but such structure is not required. Moreover, backsplicing is regulated by general splicing factors and cis-elements, but with regulatory rules distinct from canonical splicing. The resulting circRNA can be translated to generate functional proteins. Unlike linear mRNA, poly-adenosine or poly-thymidine in 3' UTR can inhibit circular mRNA translation. This study revealed that backsplicing can occur efficiently in diverse eukaryotes to generate circular mRNAs.
Keywords: alternative splicing; backsplicing; circular RNA; splicing factors; translation.
© 2015 Wang and Wang; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources