

Redox regulation of cardiomyocyte cell cycling via an ERK1/2 and c-Myc-dependent activation of cyclin D2 transcription
- PMID: 25450615
- PMCID: PMC4312357
- DOI: 10.1016/j.yjmcc.2014.10.017
Redox regulation of cardiomyocyte cell cycling via an ERK1/2 and c-Myc-dependent activation of cyclin D2 transcription
Abstract
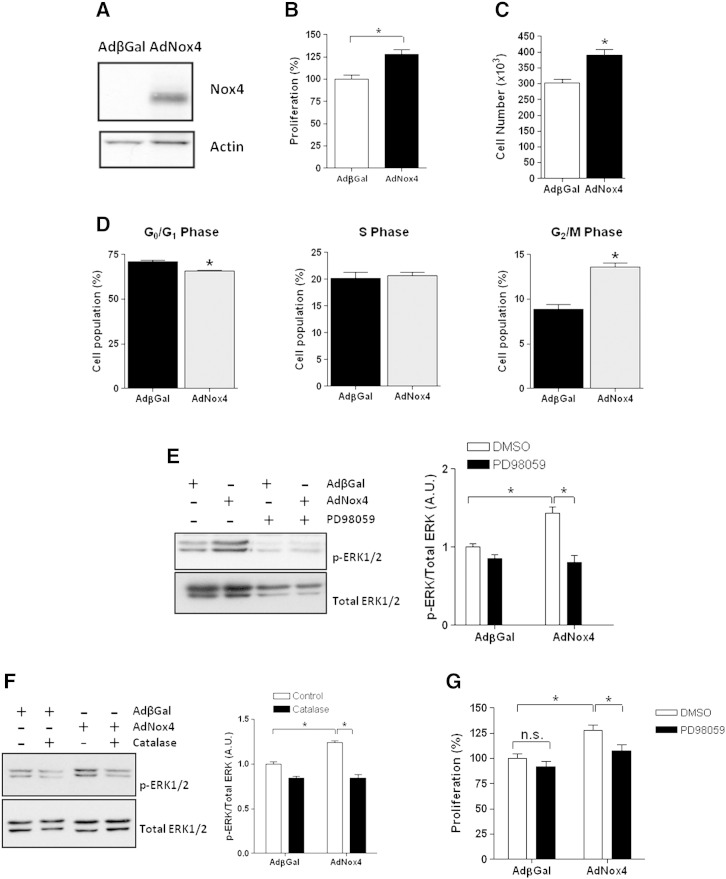
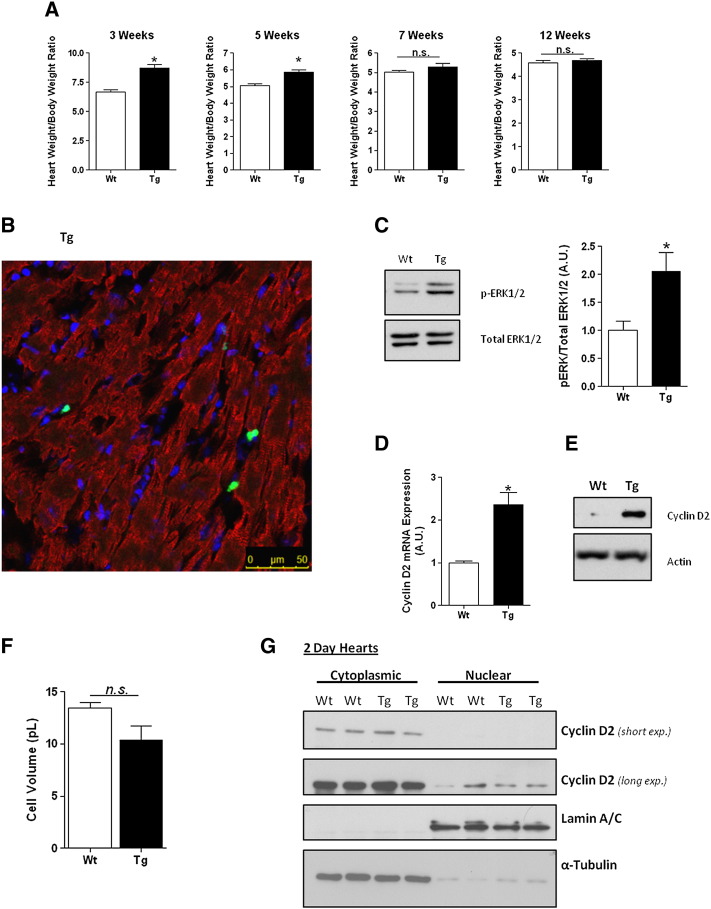
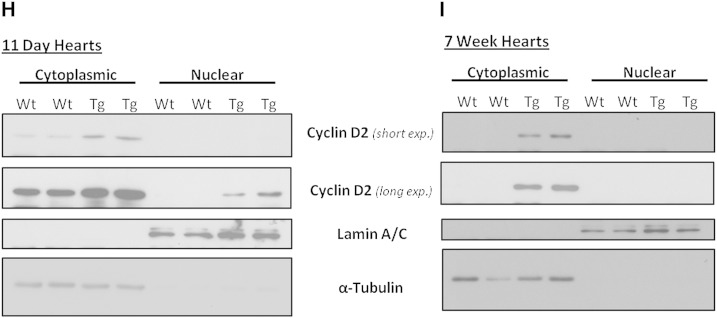
Adult mammalian cardiomyocytes have a very limited capacity to proliferate, and consequently the loss of cells after cardiac stress promotes heart failure. Recent evidence suggests that administration of hydrogen peroxide (H2O2), can regulate redox-dependent signalling pathway(s) to promote cardiomyocyte proliferation in vitro, but the potential relevance of such a pathway in vivo has not been tested. We have generated a transgenic (Tg) mouse model in which the H2O2-generating enzyme, NADPH oxidase 4 (Nox4), is overexpressed within the postnatal cardiomyocytes, and observed that the hearts of 1-3week old Tg mice pups are larger in comparison to wild type (Wt) littermate controls. We demonstrate that the cardiomyocytes of Tg mouse pups have increased cell cycling capacity in vivo as determined by incorporation of 5-bromo-2'-deoxyuridine. Further, microarray analyses of the transcriptome of these Tg mouse hearts suggested that the expression of cyclin D2 is significantly increased. We investigated the molecular mechanisms which underlie this more proliferative phenotype in isolated neonatal rat cardiomyocytes (NRCs) in vitro, and demonstrate that Nox4 overexpression mediates an H2O2-dependent activation of the ERK1/2 signalling pathway, which in turn phosphorylates and activates the transcription factor c-myc. This results in a significant increase in cyclin D2 expression, which we show to be mediated, at least in part, by cis-acting c-myc binding sites within the proximal cyclin D2 promoter. Overexpression of Nox4 in NRCs results in an increase in their proliferative capacity that is ablated by the silencing of cyclin D2. We further demonstrate activation of the ERK1/2 signalling pathway, increased phosphorylation of c-myc and significantly increased expression of cyclin D2 protein in the Nox4 Tg hearts. We suggest that this pathway acts to maintain the proliferative capacity of cardiomyocytes in Nox4 Tg pups in vivo and so delays their exit from the cell cycle after birth.
Keywords: Cardiomyocyte proliferation; Cyclin D2; ERK1/2; Nox4; Redox signalling; c-Myc.
Copyright © 2014. Published by Elsevier Ltd.
Figures







References
-
- Bicknell K.A., Coxon C.H., Brooks G. Can the cardiomyocyte cell cycle be reprogrammed? J Mol Cell Cardiol. 2007;42:706–721. - PubMed
-
- Drenckhahn J.D., Schwarz Q.P., Gray S., Laskowski A., Kiriazis H., Ming Z. Compensatory growth of healthy cardiac cells in the presence of diseased cells restores tissue homeostasis during heart development. Dev Cell. 2008;15:521–533. - PubMed
-
- Chiu J., Dawes I.W. Redox control of cell proliferation. Trends Cell Biol. 2012;22:592–601. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
