

H9c2 and HL-1 cells demonstrate distinct features of energy metabolism, mitochondrial function and sensitivity to hypoxia-reoxygenation
- PMID: 25450968
- PMCID: PMC4388199
- DOI: 10.1016/j.bbamcr.2014.11.015
H9c2 and HL-1 cells demonstrate distinct features of energy metabolism, mitochondrial function and sensitivity to hypoxia-reoxygenation
Abstract
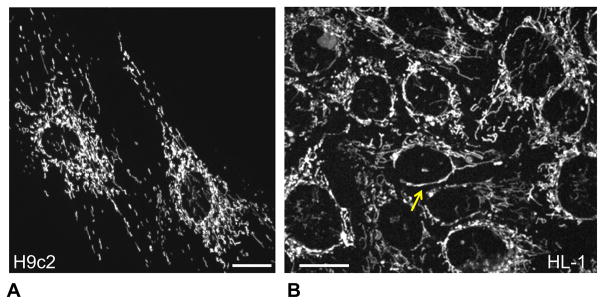
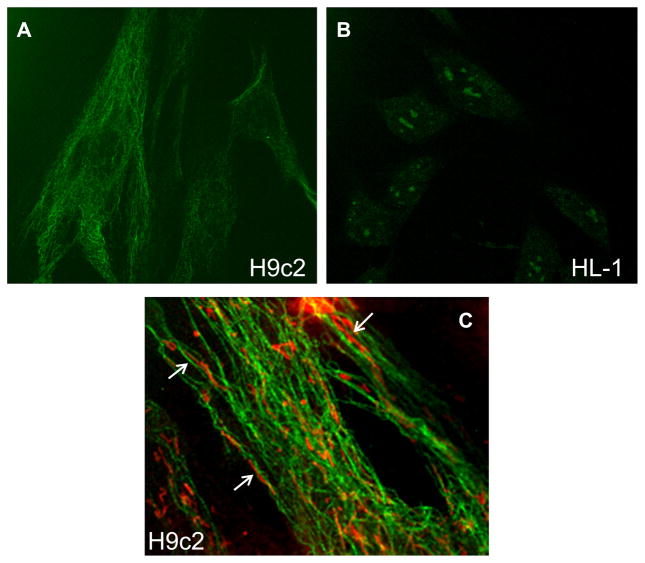
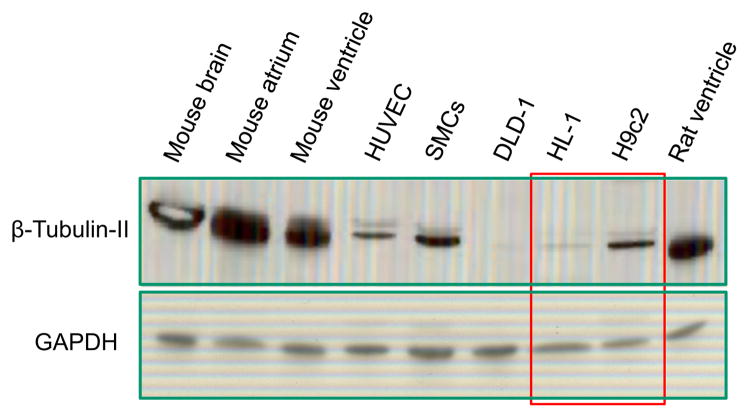
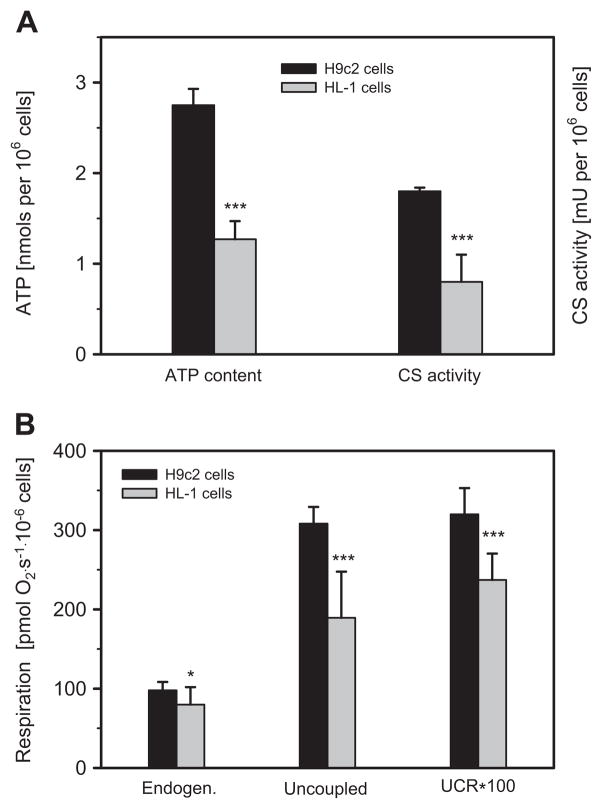
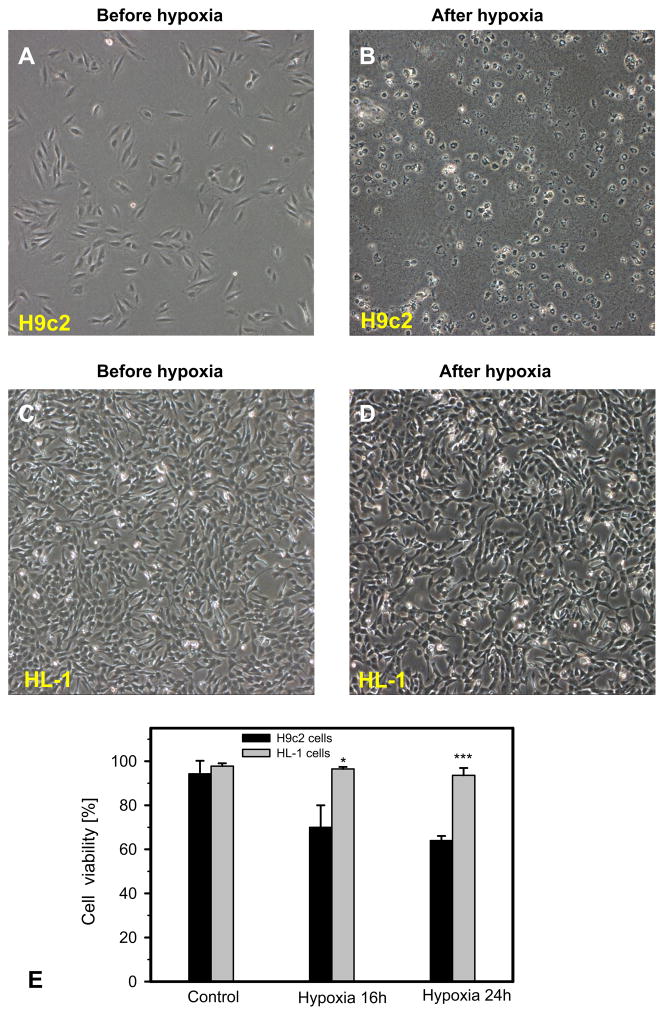
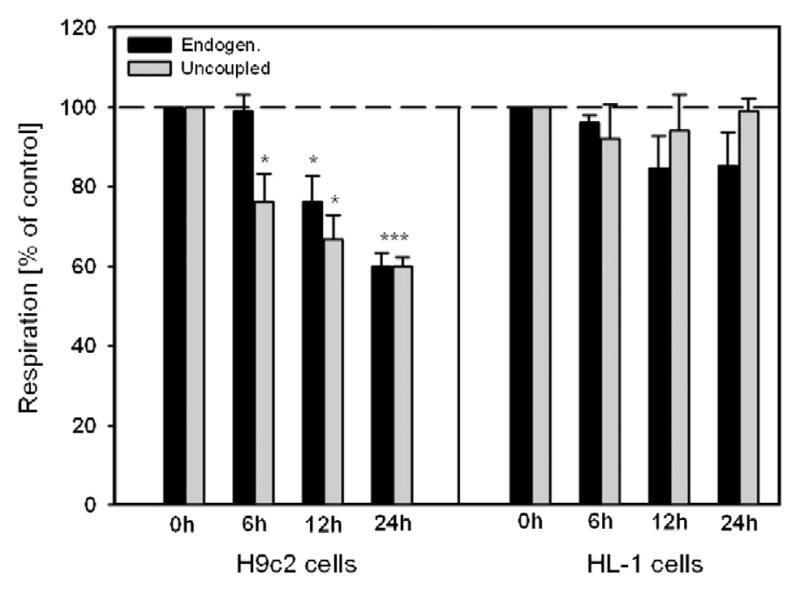
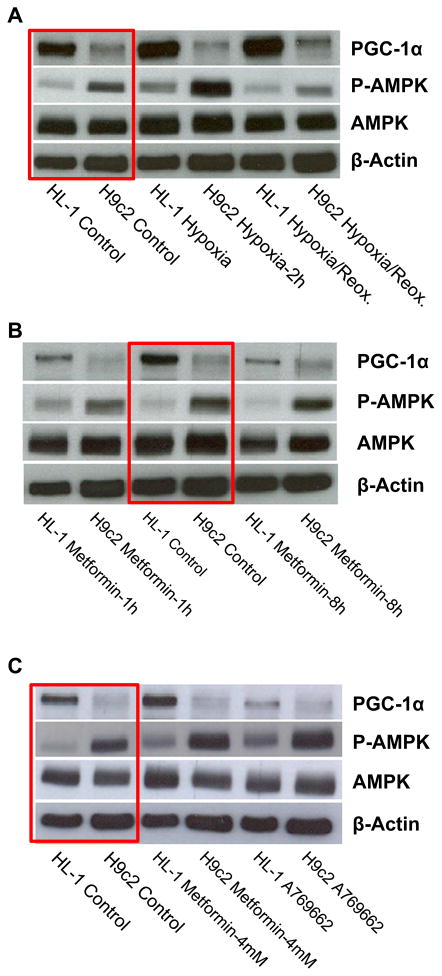
Dysfunction of cardiac energy metabolism plays a critical role in many cardiac diseases, including heart failure, myocardial infarction and ischemia-reperfusion injury and organ transplantation. The characteristics of these diseases can be elucidated in vivo, though animal-free in vitro experiments, with primary adult or neonatal cardiomyocytes, the rat ventricular H9c2 cell line or the mouse atrial HL-1 cells, providing intriguing experimental alternatives. Currently, it is not clear how H9c2 and HL-1 cells mimic the responses of primary cardiomyocytes to hypoxia and oxidative stress. In the present study, we show that H9c2 cells are more similar to primary cardiomyocytes than HL-1 cells with regard to energy metabolism patterns, such as cellular ATP levels, bioenergetics, metabolism, function and morphology of mitochondria. In contrast to HL-1, H9c2 cells possess beta-tubulin II, a mitochondrial isoform of tubulin that plays an important role in mitochondrial function and regulation. We demonstrate that H9c2 cells are significantly more sensitive to hypoxia-reoxygenation injury in terms of loss of cell viability and mitochondrial respiration, whereas HL-1 cells were more resistant to hypoxia as evidenced by their relative stability. In comparison to HL-1 cells, H9c2 cells exhibit a higher phosphorylation (activation) state of AMP-activated protein kinase, but lower peroxisome proliferator-activated receptor gamma coactivator 1-alpha levels, suggesting that each cell type is characterized by distinct regulation of mitochondrial biogenesis. Our results provide evidence that H9c2 cardiomyoblasts are more energetically similar to primary cardiomyocytes than are atrial HL-1 cells. H9c2 cells can be successfully used as an in vitro model to simulate cardiac ischemia-reperfusion injury.
Keywords: Energy metabolism; H9c2 cells; HL-1 cells; Hypoxia-reoxygenation; Mitochondria.
Copyright © 2014 Elsevier B.V. All rights reserved.
Conflict of interest statement
No confiicts of interest, financial or otherwise, are declared by the authors.
Figures
References
-
- Balaban RS, Nemoto S, Finkel T. Mitochondria, oxidants, and aging. Cell. 2005;120:483–495. - PubMed
-
- Kroemer G, Reed JC. Mitochondrial control of cell death. Nat Med. 2000;6:513–519. - PubMed
-
- Kuznetsov AV, Schneeberger S, Seiler R, Brandacher G, Mark W, Steurer W, Saks V, Usson Y, Margreiter R, Gnaiger E. Mitochondrial defects and heterogeneous cytochrome c release after cardiac cold ischemia and reperfusion. Am J Physiol Heart Circ Physiol. 2004;286:H1633–H1641. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
