

Tangled web of interactions among proteins involved in iron-sulfur cluster assembly as unraveled by NMR, SAXS, chemical crosslinking, and functional studies
- PMID: 25450980
- PMCID: PMC4390431
- DOI: 10.1016/j.bbamcr.2014.11.020
Tangled web of interactions among proteins involved in iron-sulfur cluster assembly as unraveled by NMR, SAXS, chemical crosslinking, and functional studies
Abstract
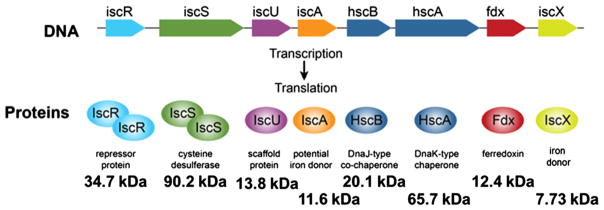
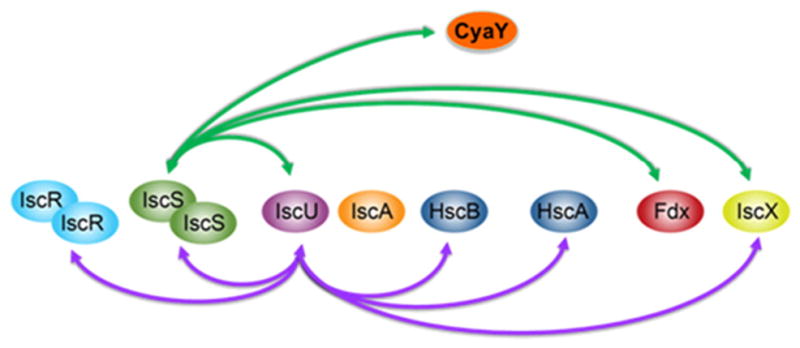
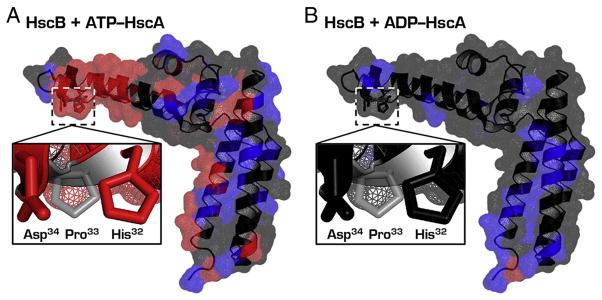
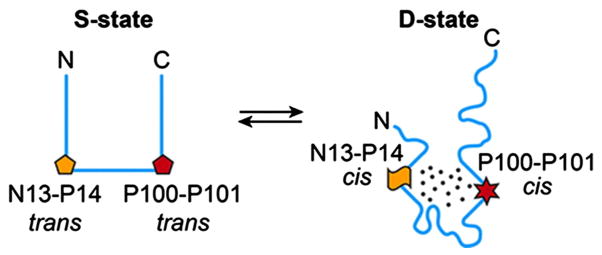
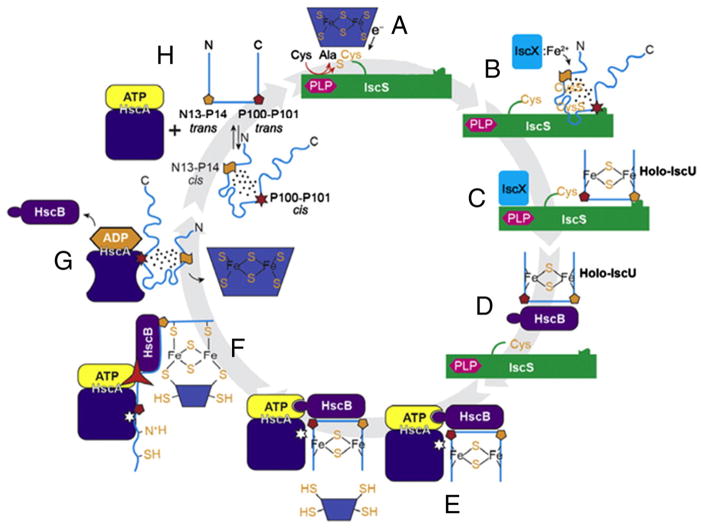
Proteins containing iron-sulfur (Fe-S) clusters arose early in evolution and are essential to life. Organisms have evolved machinery consisting of specialized proteins that operate together to assemble Fe-S clusters efficiently so as to minimize cellular exposure to their toxic constituents: iron and sulfide ions. To date, the best studied system is the iron-sulfur cluster (isc) operon of Escherichia coli, and the eight ISC proteins it encodes. Our investigations over the past five years have identified two functional conformational states for the scaffold protein (IscU) and have shown that the other ISC proteins that interact with IscU prefer to bind one conformational state or the other. From analyses of the NMR spectroscopy-derived network of interactions of ISC proteins, small-angle X-ray scattering (SAXS) data, chemical crosslinking experiments, and functional assays, we have constructed working models for Fe-S cluster assembly and delivery. Future work is needed to validate and refine what has been learned about the E. coli system and to extend these findings to the homologous Fe-S cluster biosynthetic machinery of yeast and human mitochondria. This article is part of a Special Issue entitled: Fe/S proteins: Analysis, structure, function, biogenesis and diseases.
Keywords: Conformational equilibria; Iron–sulfur cluster biosynthesis; Nuclear magnetic resonance (NMR) spectroscopy; Protein–protein interactions; Small-angle X-ray scattering (SAXS).
Copyright © 2014. Published by Elsevier B.V.
Figures
Similar articles
-
Role of IscX in iron-sulfur cluster biogenesis in Escherichia coli.J Am Chem Soc. 2014 Jun 4;136(22):7933-42. doi: 10.1021/ja501260h. Epub 2014 May 21. J Am Chem Soc. 2014. PMID: 24810328 Free PMC article.
-
Assembly of Fe/S proteins in bacterial systems: Biochemistry of the bacterial ISC system.Biochim Biophys Acta. 2015 Jun;1853(6):1436-47. doi: 10.1016/j.bbamcr.2014.12.009. Epub 2014 Dec 13. Biochim Biophys Acta. 2015. PMID: 25510311 Review.
-
Architectural Features of Human Mitochondrial Cysteine Desulfurase Complexes from Crosslinking Mass Spectrometry and Small-Angle X-Ray Scattering.Structure. 2018 Aug 7;26(8):1127-1136.e4. doi: 10.1016/j.str.2018.05.017. Epub 2018 Jul 5. Structure. 2018. PMID: 29983374 Free PMC article.
-
Molecular Mechanism of ISC Iron-Sulfur Cluster Biogenesis Revealed by High-Resolution Native Mass Spectrometry.J Am Chem Soc. 2020 Apr 1;142(13):6018-6029. doi: 10.1021/jacs.9b11454. Epub 2020 Mar 17. J Am Chem Soc. 2020. PMID: 32131593 Free PMC article.
-
Metamorphic protein IscU alternates conformations in the course of its role as the scaffold protein for iron-sulfur cluster biosynthesis and delivery.FEBS Lett. 2013 Apr 17;587(8):1172-9. doi: 10.1016/j.febslet.2013.01.003. Epub 2013 Jan 16. FEBS Lett. 2013. PMID: 23333622 Free PMC article. Review.
Cited by
-
Understanding the Molecular Basis of Multiple Mitochondrial Dysfunctions Syndrome 1 (MMDS1)-Impact of a Disease-Causing Gly208Cys Substitution on Structure and Activity of NFU1 in the Fe/S Cluster Biosynthetic Pathway.J Mol Biol. 2017 Mar 24;429(6):790-807. doi: 10.1016/j.jmb.2017.01.021. Epub 2017 Feb 1. J Mol Biol. 2017. PMID: 28161430 Free PMC article.
-
Fe-S Cluster Hsp70 Chaperones: The ATPase Cycle and Protein Interactions.Methods Enzymol. 2017;595:161-184. doi: 10.1016/bs.mie.2017.07.004. Epub 2017 Aug 21. Methods Enzymol. 2017. PMID: 28882200 Free PMC article. Review.
-
Iron-sulfur cluster biosynthesis and trafficking - impact on human disease conditions.Metallomics. 2018 Jan 24;10(1):9-29. doi: 10.1039/c7mt00180k. Metallomics. 2018. PMID: 29019354 Free PMC article. Review.
-
Cleavage and polyadenylation specificity factor 30: An RNA-binding zinc-finger protein with an unexpected 2Fe-2S cluster.Proc Natl Acad Sci U S A. 2016 Apr 26;113(17):4700-5. doi: 10.1073/pnas.1517620113. Epub 2016 Apr 11. Proc Natl Acad Sci U S A. 2016. PMID: 27071088 Free PMC article.
-
NMR as a Tool to Investigate the Processes of Mitochondrial and Cytosolic Iron-Sulfur Cluster Biosynthesis.Molecules. 2018 Aug 31;23(9):2213. doi: 10.3390/molecules23092213. Molecules. 2018. PMID: 30200358 Free PMC article. Review.
References
-
- Beinert H. A tribute to sulfur. Eur J Biochem. 2000;267:5657–5664. - PubMed
-
- Wachtershauser G. Groundworks for an evolutionary biochemistry: the iron–sulphur world. Prog Biophys Mol Biol. 1992;58:85–201. - PubMed
-
- Wachtershauser G. Origin of life. Life as we don’t know it. Science. 2000;289:1307–1308. - PubMed
-
- Huber C, Wachtershauser G. Peptides by activation of amino acids with CO on (Ni, Fe)S surfaces: implications for the origin of life. Science. 1998;281:670–672. - PubMed
-
- Cody GD, Boctor NZ, Filley TR, Hazen RM, Scott JH, Sharma A, Yoder HS., Jr Primordial carbonylated iron–sulfur compounds and the synthesis of pyruvate. Science. 2000;289:1337–1340. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P41 RR002301/RR/NCRR NIH HHS/United States
- S10RR025062/RR/NCRR NIH HHS/United States
- S10 RR025062/RR/NCRR NIH HHS/United States
- U01 GM094622/GM/NIGMS NIH HHS/United States
- S10 RR029220/RR/NCRR NIH HHS/United States
- F32 GM110939/GM/NIGMS NIH HHS/United States
- P41RR02301/RR/NCRR NIH HHS/United States
- P41GM66326/GM/NIGMS NIH HHS/United States
- S10RR023438/RR/NCRR NIH HHS/United States
- U01 GM94622/GM/NIGMS NIH HHS/United States
- P41 GM066326/GM/NIGMS NIH HHS/United States
- S10RR029220/RR/NCRR NIH HHS/United States
- S10 RR002781/RR/NCRR NIH HHS/United States
- P41 GM103399/GM/NIGMS NIH HHS/United States
- S10 RR008438/RR/NCRR NIH HHS/United States
- S10 RR023438/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
