

Ion antiport accelerates photosynthetic acclimation in fluctuating light environments
- PMID: 25451040
- PMCID: PMC4243252
- DOI: 10.1038/ncomms6439
Ion antiport accelerates photosynthetic acclimation in fluctuating light environments
Abstract
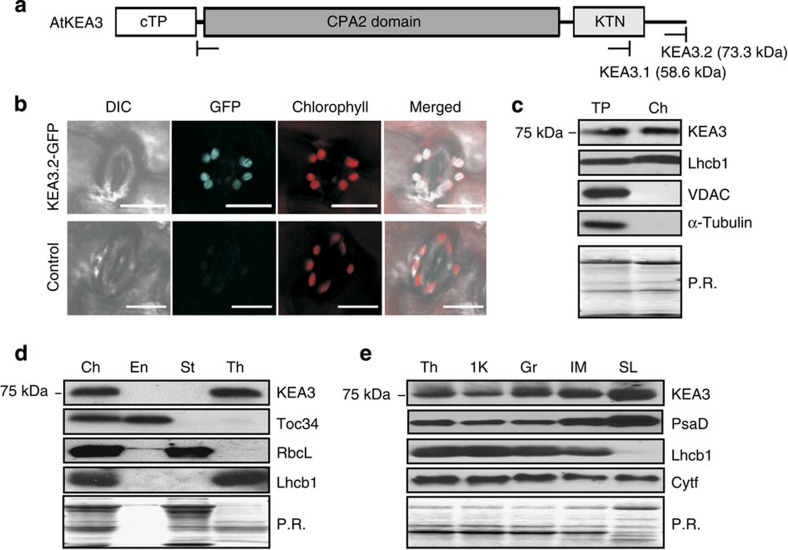
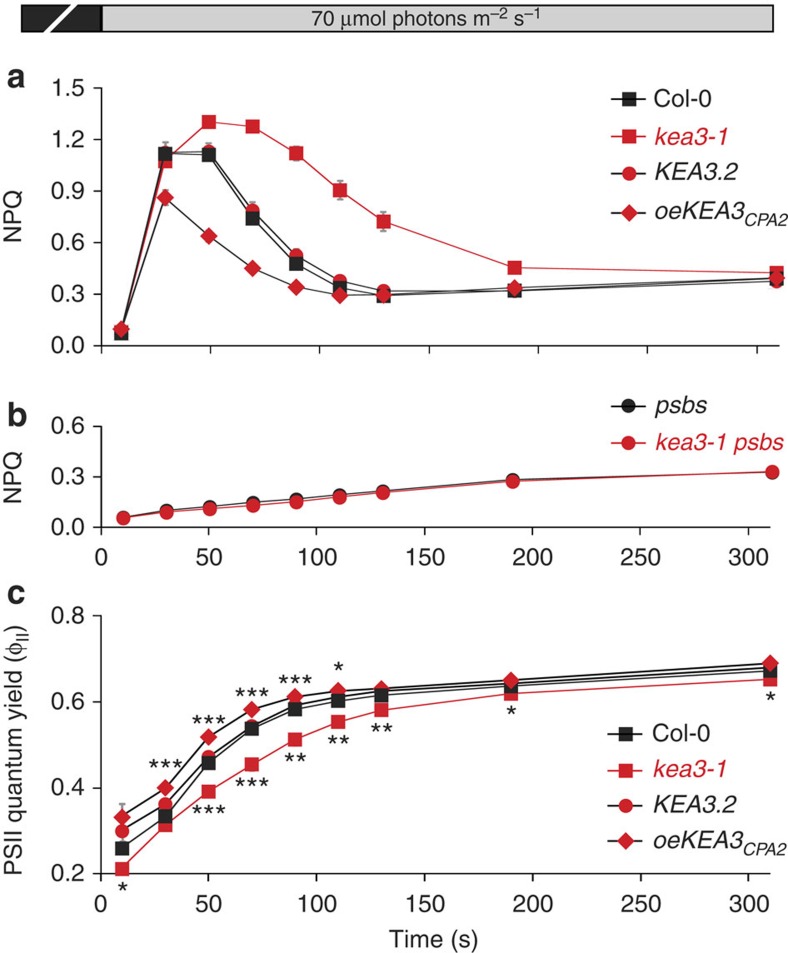
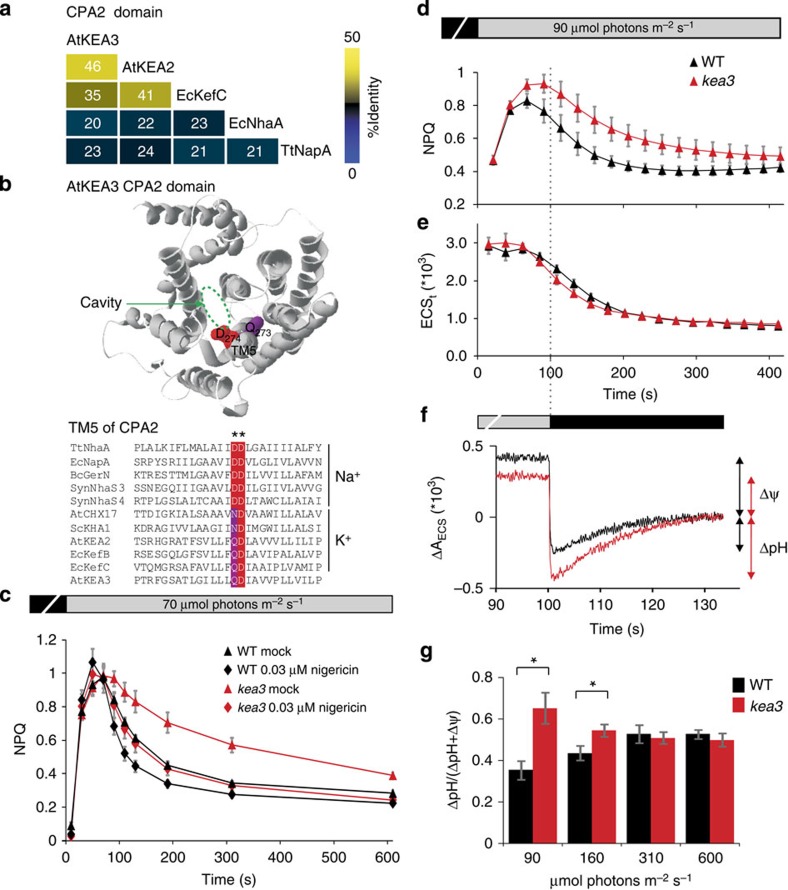
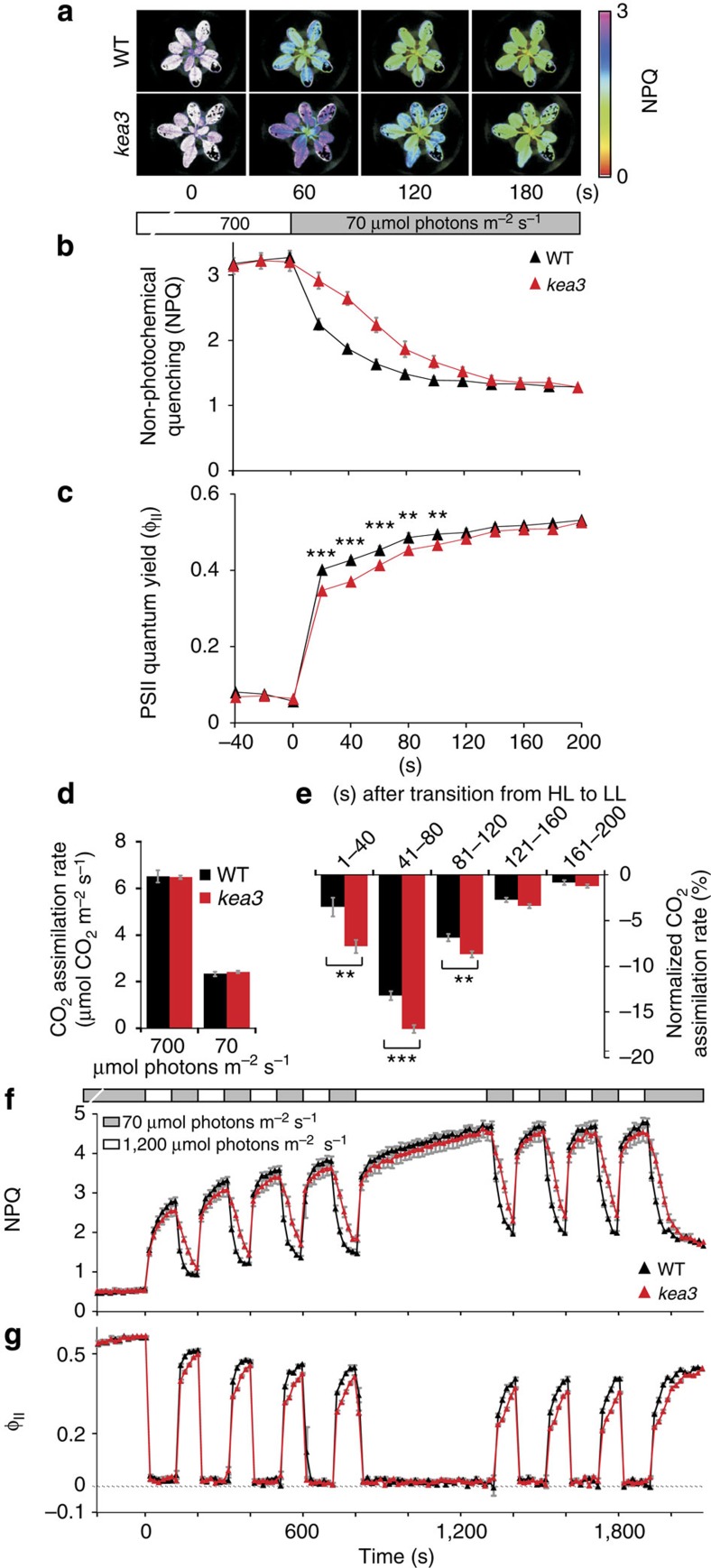
Many photosynthetic organisms globally, including crops, forests and algae, must grow in environments where the availability of light energy fluctuates dramatically. How photosynthesis maintains high efficiency despite such fluctuations in its energy source remains poorly understood. Here we show that Arabidopsis thaliana K(+) efflux antiporter (KEA3) is critical for high photosynthetic efficiency under fluctuating light. On a shift from dark to low light, or high to low light, kea3 mutants show prolonged dissipation of absorbed light energy as heat. KEA3 localizes to the thylakoid membrane, and allows proton efflux from the thylakoid lumen by proton/potassium antiport. KEA3's activity accelerates the downregulation of pH-dependent energy dissipation after transitions to low light, leading to faster recovery of high photosystem II quantum efficiency and increased CO2 assimilation. Our results reveal a mechanism that increases the efficiency of photosynthesis under fluctuating light.
Conflict of interest statement
The Carnegie Institution for Science has submitted a patent form on behalf of U.A. and M.J. on aspects of the findings.
Figures
References
-
- Pearcy R. W. Sunflecks and photosynthesis in plant canopies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 41, 421–453 (1990).
-
- Reinhard K., Smith W. K. & Carter G. A. Clouds and cloud immersion alter photosynthetic light quality in a temperate mountain cloud forest. Botany 88, 462–470 (2010).
-
- Kramer D. M., Cruz J. A. & Kanazawa A. Balancing the central roles of the thylakoid proton gradient. Trends Plant. Sci. 8, 27–32 (2003). - PubMed
-
- Allen J. Photosynthesis of ATP-electrons, proton pumps, rotors, and poise. Cell 110, 273–276 (2002). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
