

The avian coronavirus spike protein
- PMID: 25451062
- PMCID: PMC7114429
- DOI: 10.1016/j.virusres.2014.10.009
The avian coronavirus spike protein
Abstract
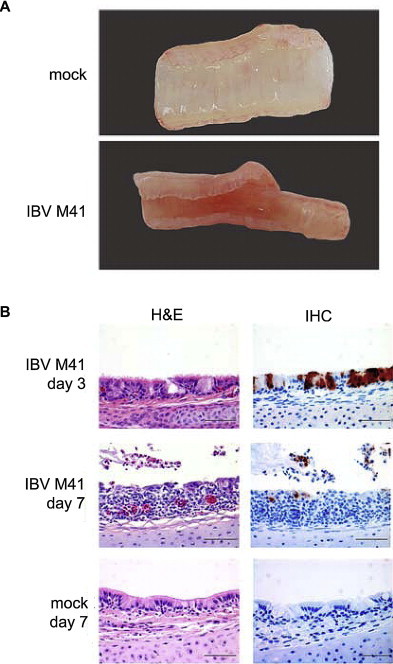
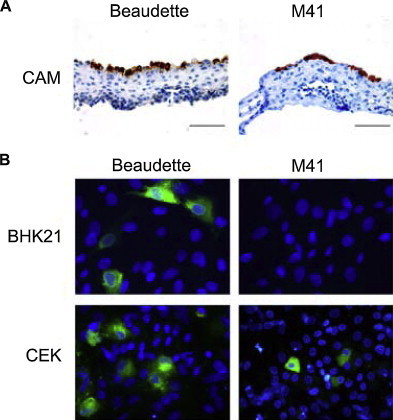
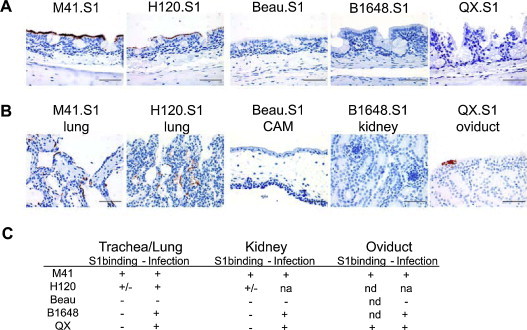
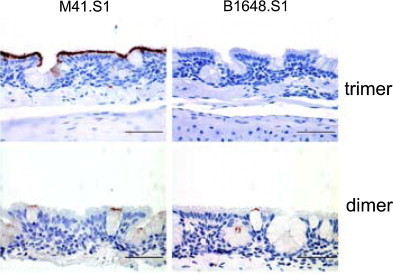
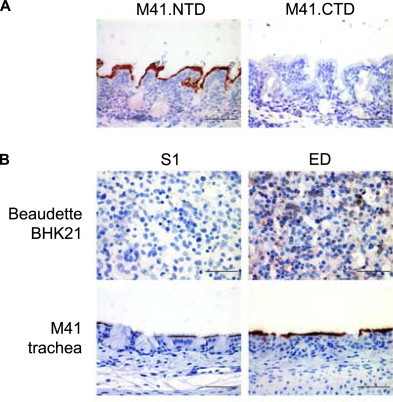
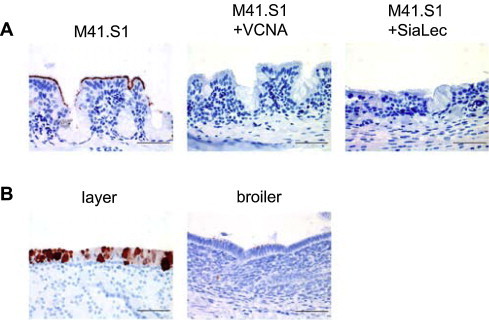
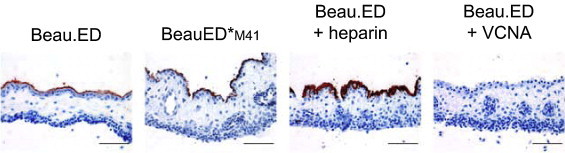
Avian coronaviruses of the genus Gammacoronavirus are represented by infectious bronchitis virus (IBV), the coronavirus of chicken. IBV causes a highly contagious disease affecting the respiratory tract and, depending on the strain, other tissues including the reproductive and urogenital tract. The control of IBV in the field is hampered by the many different strains circulating worldwide and the limited protection across strains due to serotype diversity. This diversity is believed to be due to the amino acid variation in the S1 domain of the major viral attachment protein spike. In the last years, much effort has been undertaken to address the role of the avian coronavirus spike protein in the various steps of the virus' live cycle. Various models have successfully been developed to elucidate the contribution of the spike in binding of the virus to cells, entry of cell culture cells and organ explants, and the in vivo tropism and pathogenesis. This review will give an overview of the literature on avian coronavirus spike proteins with particular focus on our recent studies on binding of recombinant soluble spike protein to chicken tissues. With this, we aim to summarize the current understanding on the avian coronavirus spike's contribution to host and tissue predilections, pathogenesis, as well as its role in therapeutic and protective interventions.
Keywords: Avian coronavirus; Binding; Gammacoronavirus; Infectious bronchitis virus IBV; Spike; Tropism.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
-
- Abd El Rahman S., El-Kenawy A.A., Neumann U., Herrler G., Winter C. Comparative analysis of the sialic acid binding activity and the tropism for the respiratory epithelium of four different strains of avian infectious bronchitis virus. Avian Pathol. 2009;38:41–45. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
