

Theta-burst LTP
- PMID: 25452022
- PMCID: PMC4411212
- DOI: 10.1016/j.brainres.2014.10.034
Theta-burst LTP
Abstract
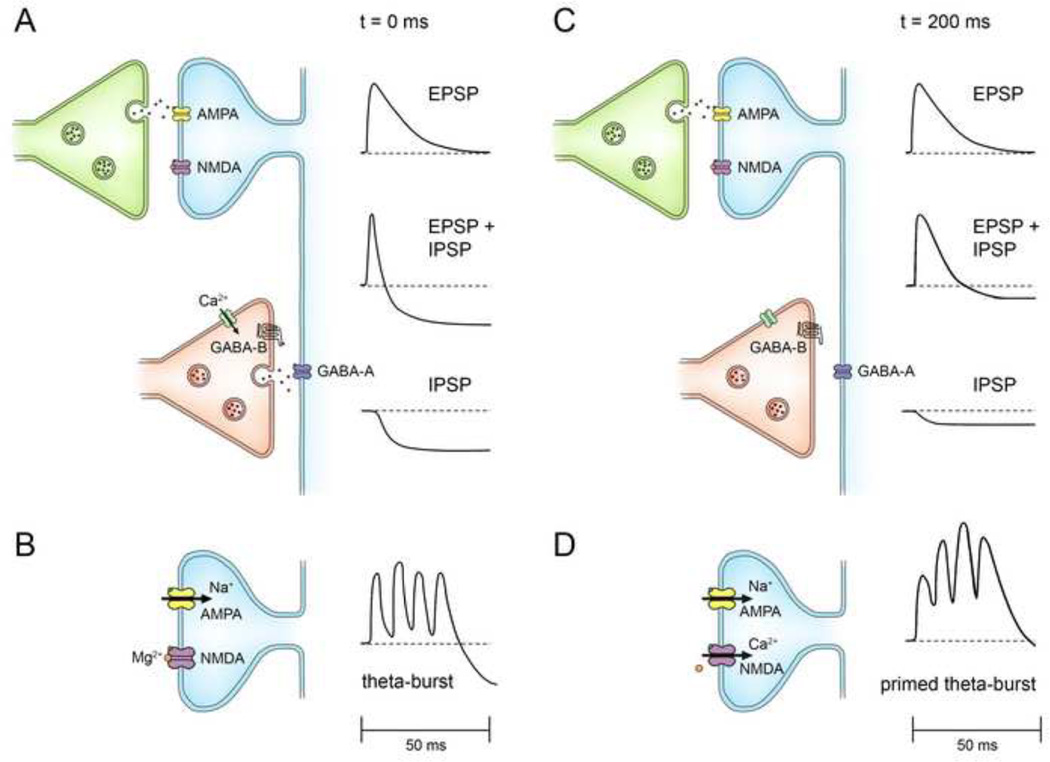
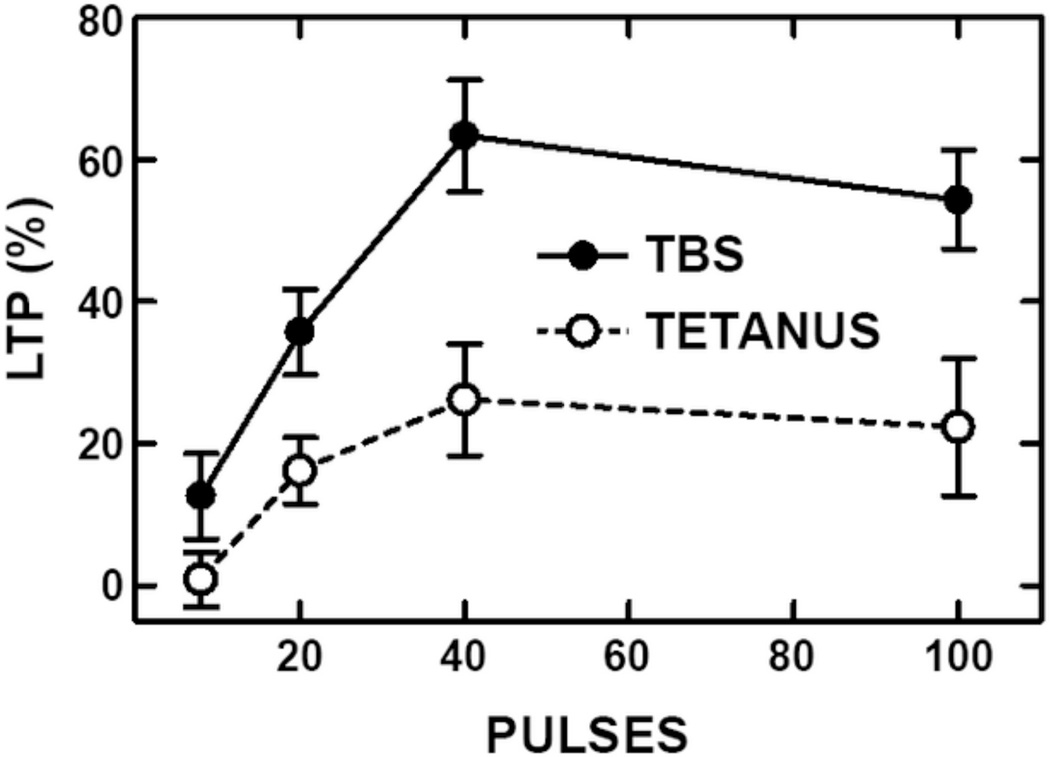
This review covers the spatial and temporal rules governing induction of hippocampal long-term potentiation (LTP) by theta-burst stimulation. Induction of LTP in field CA1 by high frequency stimulation bursts that resemble the burst discharges (complex-spikes) of hippocampal pyramidal neurons involves a multiple-step mechanism. A single burst is insufficient for LTP induction because it evokes both excitatory and inhibitory currents that partially cancel and limit postsynaptic depolarization. Bursts repeated at the frequency (~5 Hz) of the endogenous theta rhythm induce maximal LTP, primarily because this frequency disables feed-forward inhibition and allows sufficient postsynaptic depolarization to activate voltage-sensitive NMDA receptors. The disinhibitory process, referred to as "priming", involves presynaptic GABA autoreceptors that inhibit GABA release. Activation of NMDA receptors allows a calcium flux into dendritic spines that serves as the proximal trigger for LTP. We include new data showing that theta-burst stimulation is more efficient than other forms of stimulation for LTP induction. In addition, we demonstrate that associative interactions between synapses activated during theta-bursts are limited to major dendritic domains since such interactions occur within apical or basal dendritic trees but not between them. We review evidence that recordings of electrophysiological responses during theta burst stimulation can help to determine if experimental manipulations that affect LTP do so by affecting events antecedent to the induction process, such as NMDA receptor activation, or downstream signaling cascades that result from postsynaptic calcium fluxes. Finally, we argue that theta-burst LTP represents a minimal model for stable, non-decremental LTP that is more sensitive to a variety of experimental manipulations than is LTP induced by other stimulation paradigms. This article is part of a Special Issue entitled SI: Brain and Memory.
Keywords: AMPA; CA1; GABA; Hippocampus; LTP; Long-term potentiation; NMDA; Theta burst stimulation.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures




Similar articles
-
LTP in hippocampal area CA1 is induced by burst stimulation over a broad frequency range centered around delta.Learn Mem. 2009 Jan 7;16(1):69-81. doi: 10.1101/lm.1179109. Print 2009 Jan. Learn Mem. 2009. PMID: 19144965 Free PMC article.
-
Role of N-methyl-D-aspartate receptors in the induction of synaptic potentiation by burst stimulation patterned after the hippocampal theta-rhythm.Brain Res. 1988 Feb 16;441(1-2):111-8. doi: 10.1016/0006-8993(88)91388-1. Brain Res. 1988. PMID: 2896049
-
Behavioral Timescale Cooperativity and Competitive Synaptic Interactions Regulate the Induction of Complex Spike Burst-Dependent Long-Term Potentiation.J Neurosci. 2022 Mar 30;42(13):2647-2661. doi: 10.1523/JNEUROSCI.1950-21.2022. Epub 2022 Feb 8. J Neurosci. 2022. PMID: 35135856 Free PMC article.
-
Phasic modulation of hippocampal synaptic plasticity by theta rhythm.Behav Neurosci. 2020 Dec;134(6):595-612. doi: 10.1037/bne0000354. Epub 2020 Jan 9. Behav Neurosci. 2020. PMID: 31916794 Review.
-
Muscarinic Receptors, from Synaptic Plasticity to its Role in Network Activity.Neuroscience. 2021 Feb 21;456:60-70. doi: 10.1016/j.neuroscience.2020.04.005. Epub 2020 Apr 8. Neuroscience. 2021. PMID: 32278062 Review.
Cited by
-
Noninvasive Brain Stimulation & Space Exploration: Opportunities and Challenges.Neurosci Biobehav Rev. 2020 Dec;119:294-319. doi: 10.1016/j.neubiorev.2020.09.005. Epub 2020 Sep 13. Neurosci Biobehav Rev. 2020. PMID: 32937115 Free PMC article. Review.
-
Ripple-triggered stimulation of the locus coeruleus during post-learning sleep disrupts ripple/spindle coupling and impairs memory consolidation.Learn Mem. 2016 Apr 15;23(5):238-48. doi: 10.1101/lm.040923.115. Print 2016 May. Learn Mem. 2016. PMID: 27084931 Free PMC article.
-
Apoptosis signal-regulating kinase 1 (Ask1) deficiency alleviates MPP+-induced impairment of evoked dopamine release in the mouse hippocampus.Front Cell Neurosci. 2024 Feb 13;18:1288991. doi: 10.3389/fncel.2024.1288991. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38414754 Free PMC article.
-
The α2δ-1-NMDA receptor coupling is essential for corticostriatal long-term potentiation and is involved in learning and memory.J Biol Chem. 2018 Dec 14;293(50):19354-19364. doi: 10.1074/jbc.RA118.003977. Epub 2018 Oct 24. J Biol Chem. 2018. PMID: 30355732 Free PMC article.
-
Effects of theta phase precessing optogenetic intervention on hippocampal neuronal reactivation and spatial maps.iScience. 2023 Jun 28;26(7):107233. doi: 10.1016/j.isci.2023.107233. eCollection 2023 Jul 21. iScience. 2023. PMID: 37534136 Free PMC article.
References
-
- Andersen P. Foreword: LTP-an exciting and continuing saga. In: Baudry M, Davis JL, editors. In: Long-Term Potentiation: A Debate of Current Issues. Cambridge, MA: MIT Press; 1991. pp. xiii–xvii. Vol.
-
- Arai A, Lynch G. Factors regulating the magnitude of long-term potentiation induced by theta pattern stimulation. Brain Res. 1992;598:173–184. - PubMed
-
- Arai A, Silberg J, Lynch G. Differences in the refractory properties of two distinct inhibitory circuitries in field CA1 of the hippocampus. Brain Res. 1995;704:298–306. - PubMed
-
- Arai AC, Xia YF, Suzuki E. Modulation of AMPA receptor kinetics differentially influences synaptic plasticity in the hippocampus. Neuroscience. 2004;123:1011–1024. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
