

Nedd8 regulates inflammasome-dependent caspase-1 activation
- PMID: 25452302
- PMCID: PMC4285429
- DOI: 10.1128/MCB.00775-14
Nedd8 regulates inflammasome-dependent caspase-1 activation
Abstract
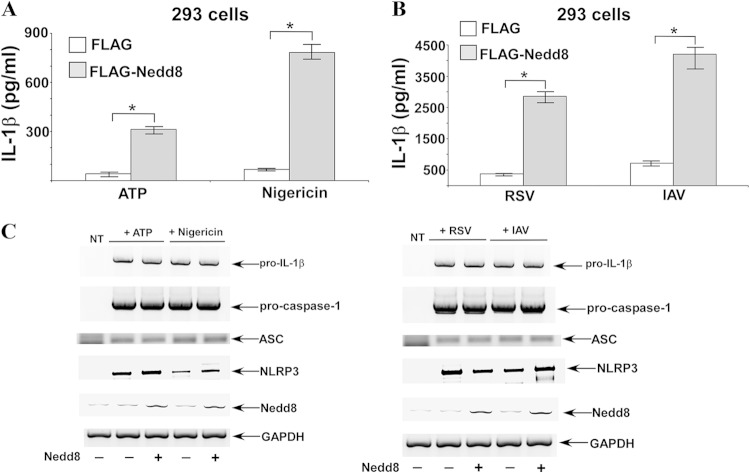
Caspase-1 is activated by the inflammasome complex to process cytokines like interleukin-1β (IL-1β). Pro-caspase-1 consists of three domains, CARD, p20, and p10. Association of pro-caspase-1 with the inflammasome results in initiation of its autocatalytic activity, culminating in self-cleavage that generates catalytically active subunits (p10 and p20). In the current study, we show that Nedd8 is required for efficient self-cleavage of pro-caspase-1 to generate its catalytically active subunits. Nedd8 silencing or treating cells with the neddylation inhibitor MLN4924 led to diminished caspase-1 processing and reduced IL-1β maturation following inflammasome activation. Coimmunoprecipitation and mass spectrometric analysis of 293 cells overexpressing pro-caspase-1 (and CARD) and Nedd8 suggested possible neddylation of caspase-1 CARD. Following inflammasome activation in primary macrophages, we observed colocalization of endogenous Nedd8 with caspase-1. Similarly, interaction of endogenous Nedd8 with caspase-1 CARD was detected in inflammasome-activated macrophages. Furthermore, enhanced autocatalytic activity of pro-caspase-1 was observed following Nedd8 overexpression in 293 cells, and such activity in inflammasome-activated macrophages was drastically diminished upon treatment of cells with MLN4924. Thus, our studies demonstrate a role of Nedd8 in regulating caspase-1 activation following inflammasome activation, presumably via augmenting autoprocessing/cleavage of pro-caspase-1 into its corresponding catalytically active subunits.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures








References
-
- Thornberry NA, Bull HG, Calaycay JR, Chapman KT, Howard AD, Kostura MJ, Miller DK, Molineaux SM, Weidner JR, Aunins J, Elliston KO, Ayala JM, Casano FJ, Chin J, Ding GJ-F, Egger LA, Gaffney EP, Limjuco G, Palyha OC, Raju SM, Rolando AM, Salley JP, Yamin T-T, Lee TD, Shively JE, MacCross M, Mumford RA, Schmidt JA, Tocci MJ. 1992. A novel heterodimeric cysteine protease is required for interleukin-1 beta processing in monocytes. Nature 356:768–774. doi: 10.1038/356768a0. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI083387/AI/NIAID NIH HHS/United States
- T32 DE014318/DE/NIDCR NIH HHS/United States
- 7P30CA54174-14/CA/NCI NIH HHS/United States
- P01 AG019316/AG/NIA NIH HHS/United States
- P30 AG013319/AG/NIA NIH HHS/United States
- NIH-NIA P01AG19316/AG/NIA NIH HHS/United States
- DE14318/DE/NIDCR NIH HHS/United States
- NIH-NIA P30AG013319/AG/NIA NIH HHS/United States
- UL1 TR000149/TR/NCATS NIH HHS/United States
- P30 CA054174/CA/NCI NIH HHS/United States
- UL1 TR001120/TR/NCATS NIH HHS/United States
- NIH-NCI P30 CA54174/CA/NCI NIH HHS/United States
- 8UL1 TR000149/TR/NCATS NIH HHS/United States
- AI083387/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous