

Review
doi: 10.1101/cshperspect.a019315.
Epigenetic regulation in plants
Affiliations
- PMID: 25452385
- PMCID: PMC4292151
- DOI: 10.1101/cshperspect.a019315
Item in Clipboard
Review
Epigenetic regulation in plants
Cold Spring Harb Perspect Biol.
.
Abstract
The study of epigenetics in plants has a long and rich history, from initial descriptions of non-Mendelian gene behaviors to seminal discoveries of chromatin-modifying proteins and RNAs that mediate gene silencing in most eukaryotes, including humans. Genetic screens in the model plant Arabidopsis have been particularly rewarding, identifying more than 130 epigenetic regulators thus far. The diversity of epigenetic pathways in plants is remarkable, presumably contributing to the phenotypic plasticity of plant postembryonic development and the ability to survive and reproduce in unpredictable environments.
Copyright © 2014 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

Unique aspects of the plant life cycle. Plants can propagate sexually (gametogenesis, fertilization, and seed formation; right) as well as somatically (vegetative explants, cell de- and re-differentiation, or somatic embryogenesis; left). The body of higher plants with roots, stem, leaves, and flowers is the diploid sporophyte. During meiosis, the chromosome number is reduced by half. Whereas in animals the meiotic products become the gametes without further division and fuse directly to produce the diploid embryo, plants form haploid male or female gametophytes via two or three mitotic divisions to form pollen or embryo sacs, respectively. The pollen grain ultimately contains one vegetative (white) and two generative (black) nuclei. The two generative nuclei fertilize the egg cell (black) and the central cell, the latter having a diploid nucleus derived from fusion of the two polar nuclei (yellow). This double fertilization gives rise to a diploid embryo and a triploid endosperm, the latter providing a nutrient source for the developing embryo. After seed germination, the embryo will grow into a new sporophyte. Most plants also have the potential for vegetative propagation through activation of lateral meristems, outgrowth of specialized organs (tubers, rhizomes or stolons), amplification in tissue culture, or regeneration from individual somatic cells after removal of the cell wall (protoplasts). Endoreduplication is frequent in plants, producing polyploid cells or tissues. Plants can be grafted to produce chimeras.

Assays for epigenetic control in plants. Genes determining coloration of plant tissue allow easy and inexpensive gene expression readout in vivo. (A) Expression of the dihydroflavonol reductase (DFR) gene is required for dark purple petunia flowers, whereas silencing of the DFR promoter gives rise to variegated, light coloration. (B) Seeds from Arabidopsis expressing the chalcone synthase (CHS) gene have dark seed coats, whereas silencing of CHS upon expression of a homologous transgene results in yellow seeds. (C) Maize plants with the B-I gene have purple pigmentation, in contrast to green plants with a paramutagenic, inactive B′ allele whose DNA sequence is identical. (D) Maize ear, segregating a transposon insertion (Spm) in the B-Peru gene required for anthocyanin pigment. Purple kernels represent revertants in which the Spm element excised from the gene in the germline. The heavily spotted kernels contain the Spm element in the active form that induces frequent somatic excision sectors during kernel development. The kernels with rare, small purple sectors represent kernels in which the Spm element has been epigenetically silenced. (E) The dark color of soybeans (middle) is extinguished in cultivated varieties (left) because of natural posttranscriptional silencing of the CHS gene and can be partially reversed by infection of the parental plant with a virus possessing a PTGS suppressor protein, producing a mottled pattern (right). (F) Epigenetic regulation can become manifest also in plant morphology; reduced function of a chromatin assembly factor subunit leads to a “fasciated” Arabidopsis stem. (G) Release of silencing from a transgenic resistance marker in Arabidopsis can be scored by growth on selective medium. (A, Courtesy of Jan Kooter; B, courtesy of Ian Furner; C, reprinted from Chandler et al. 2000, with kind permission from Springer Science and Business Media; D, courtesy of Vicki Chandler; E, reprinted, with permission, from Senda et al. 2004, © American Society of Plant Biologists.)
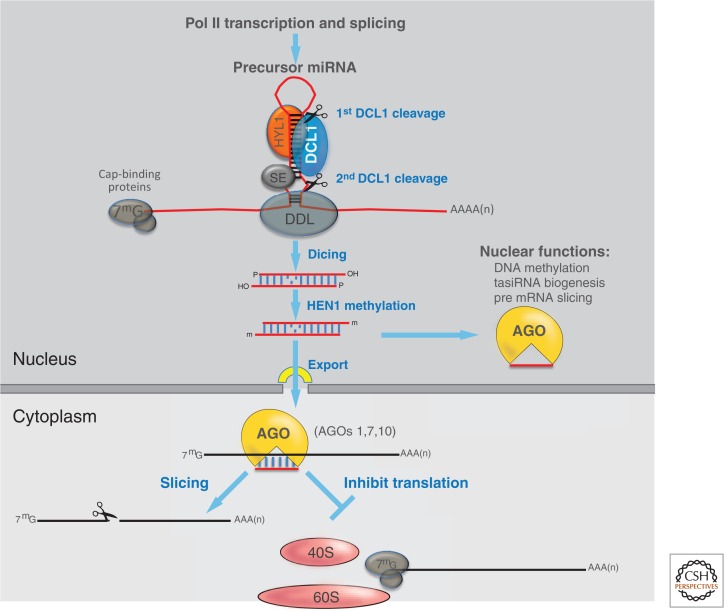
miRNA biogenesis and modes of action. Capped and polyadenylated transcripts of nuclear RNA Pol II that form imperfect hairpins serve as the precursors for miRNAs. DCL1 cleaves these precursors with the help of the HYL1, DDL1, and SE nucleic acid–binding proteins. Diced products are end-methylated by HEN1 and exported to the cytosplasm in a process involving the exportin 5 homolog, HST. In association with AGO1 or a related AGO protein, such as AGO10, the miRNA guides the cleavage or translational inhibition of complementary mRNAs. Nuclear functions in which miRNAs are implicated include specific cases of cytosine methylation, trans-acting siRNA (tasiRNA) production or pre-mRNA degradation.

tasiRNA biogenesis and function. Specific miRNAs, associated with AGO1 or AGO7, target and cleave the transcripts of TAS gene loci, ultimately giving rise to tasiRNAs. In the case of AGO1, initiating miRNAs that are 22 nt in length induce the recruitment of the RNA-dependent RNA polymerase, RDR6 (and its partner SGS3), to the 3′ cleaved fragment resulting in transcription of the complementary strand to generate dsRNA. Dicing by DCL4, aided by the dsRNA-binding protein, DRB4, results in the production of 21-nt tasiRNAs that are phased with respect to the miRNA-cleaved end. These tasiRNAs, in turn, target specific complementary RNAs in association with AGO1, thus behaving like target-specific miRNAs.

PTGS induced by viruses or transgenes. Double-stranded precursors of siRNAs that bring about PTGS can be dsRNA viral genomes or replication intermediates, dsRNAs formed by overlapping transcripts from adjacent transcription units, or inverted repeats resulting from tandem transgenes that integrate in opposite orientations. Alternatively, single-stranded RNAs can be made double-stranded by the action of RNA-dependent RNA polymerases, such as RDR6 or RDR1. Dicing can then occur by DCLs 2, -3 or -4, with 22-nt DCL2 products and 21-nt DCL4 products being primarily associated with PTGS. On association with AGO1, the siRNAs target complementary RNAs for degradation or translational inhibition.

RdDM. Retrotransposons, viruses, transgenes, or repetitive genes are subject to silencing by RdDM. RNA Pol IV is thought to generate single-stranded RNAs (bottom left) that serve as templates for the RNA-dependent RNA polymerase, RDR2. Resulting dsRNAs are diced by DCL3 to generate 24-nt siRNA duplexes that are then methylated by HEN1 and loaded onto AGO4, or its closely related family members, AGO6 or AGO9. dsRNA derived from inverted repeat transgenes or viruses (top left) can bypass the need for RNA Pol IV and RDR2. AGO-siRNA complexes are recruited to their sites of action by binding to transcripts generated by RNA Pol V, as well as by physical interactions with the carboxy-terminal domain of the RNA Pol V largest subunit. At some loci, RNA Pol II is thought to substitute for RNA Pol V for the production of scaffold transcripts to which AGO-siRNA complexes bind. The DDR complex (DRD1, DMS3, and RDM1) enables RNA Pol V transcription. The RDM1 subunit of the DDR complex also interacts with AGO4 and the de novo cytosine methyltransferase, DRM2, thus potentially serving as a bridge that recruits DRM2 to sites of RNA Pol V transcription.

Transitivity: amplification and spreading of secondary siRNAs from the site of a primary siRNA target. Primary siRNAs are thought to prime RNA-dependent RNA polymerase activity, resulting in dsRNAs extending toward the 5′ end of the target RNA (left). Subsequent dicing gives rise to secondary siRNAs in the region upstream of the initiating primary siRNA. 22-nt siRNAs also have the property of recruiting RDR6 to the 3′ fragment following AGO slicing (right), resulting in dsRNA and secondary siRNA biogenesis in the region downstream from the initiating, primary siRNA.
References
WWW RESOURCES
-
- http://www.arabidopsis.leeds.ac.uk/act/coexpanalyser.php Arabidopsis coexpression mining.
-
- http://asrp.cgrb.oregonstate.edu Arabidopsis small RNA.
-
- http://bbc.botany.utoronto.ca/efp/cgi-bin/efpWeb.cgi Gene expression in Arabidopsis.
-
- http://www.chromdb.org Chromatin genes.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources