

Iron-binding E3 ligase mediates iron response in plants by targeting basic helix-loop-helix transcription factors
- PMID: 25452667
- PMCID: PMC4281009
- DOI: 10.1104/pp.114.250837
Iron-binding E3 ligase mediates iron response in plants by targeting basic helix-loop-helix transcription factors
Abstract
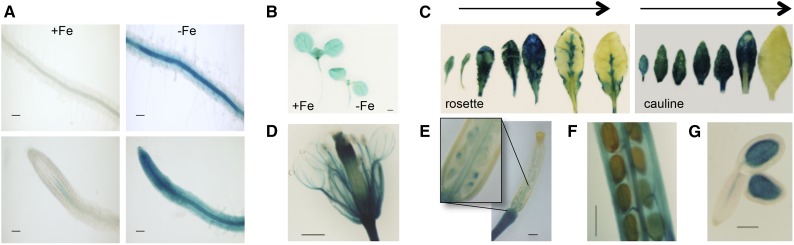
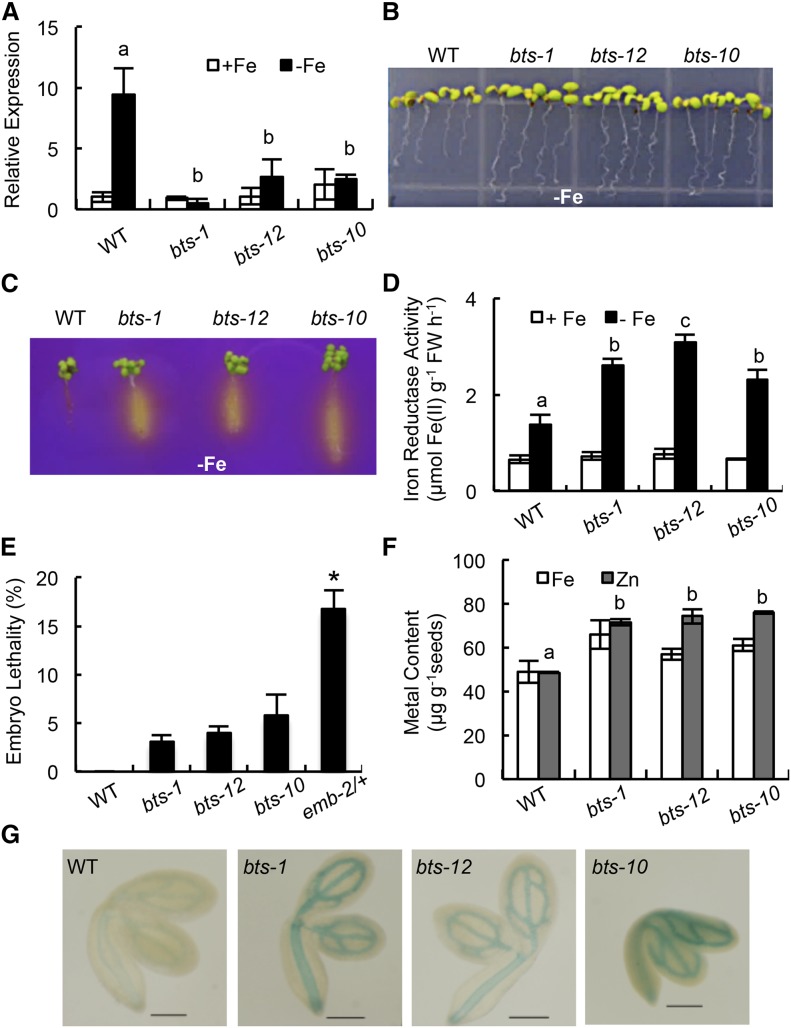
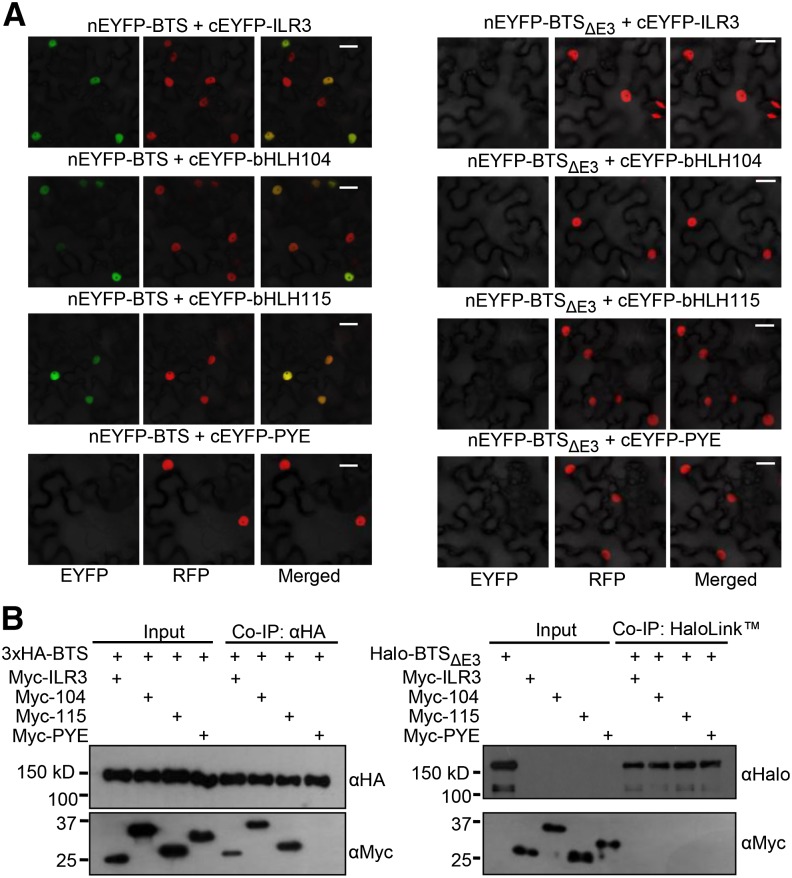
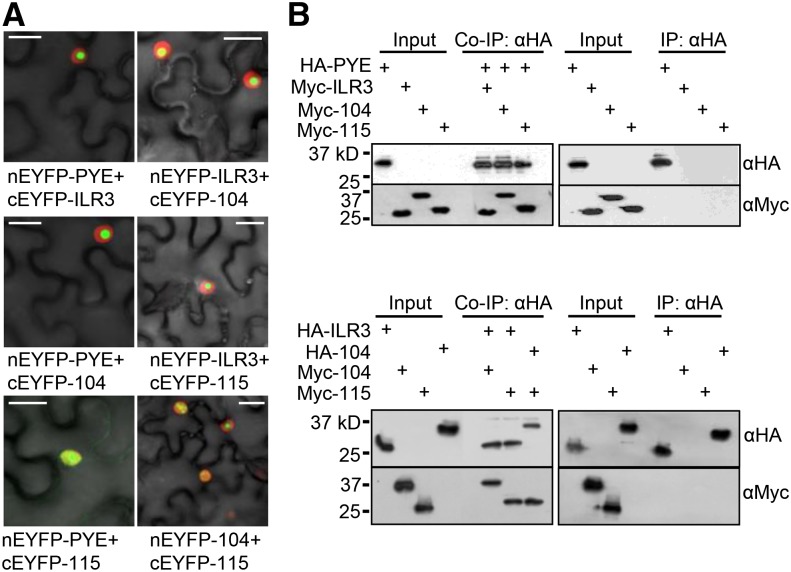
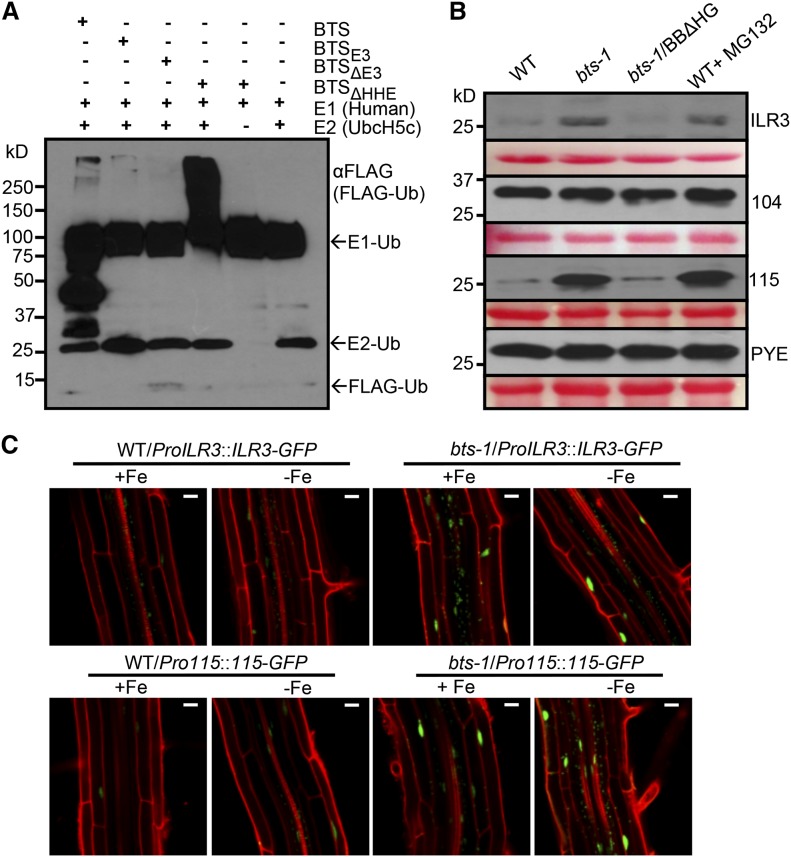
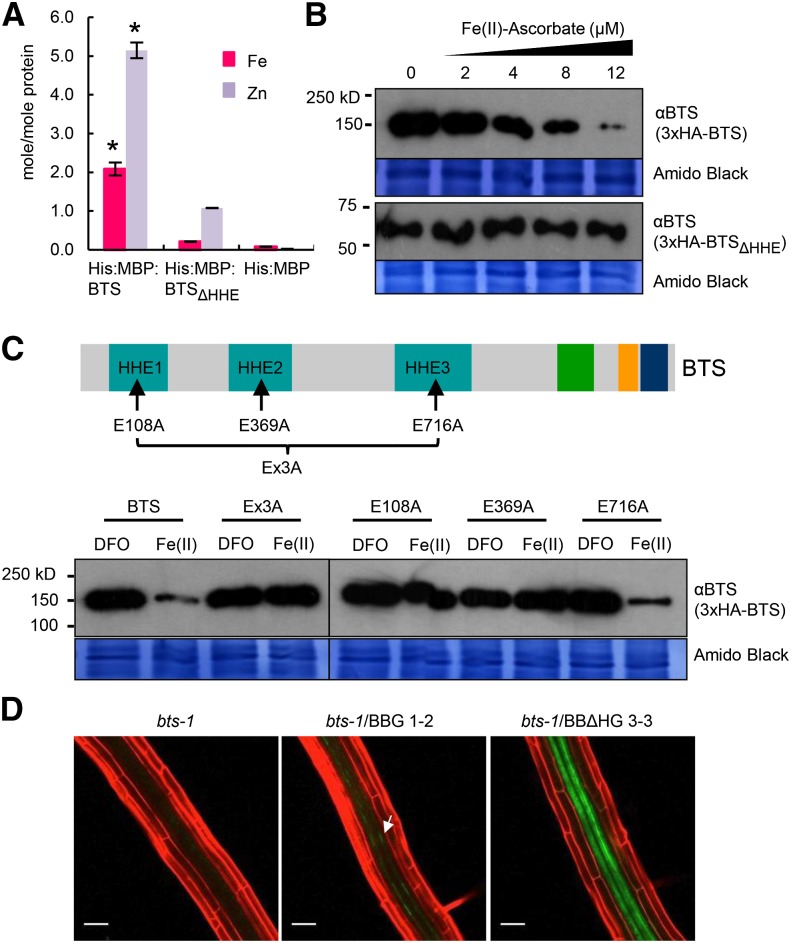
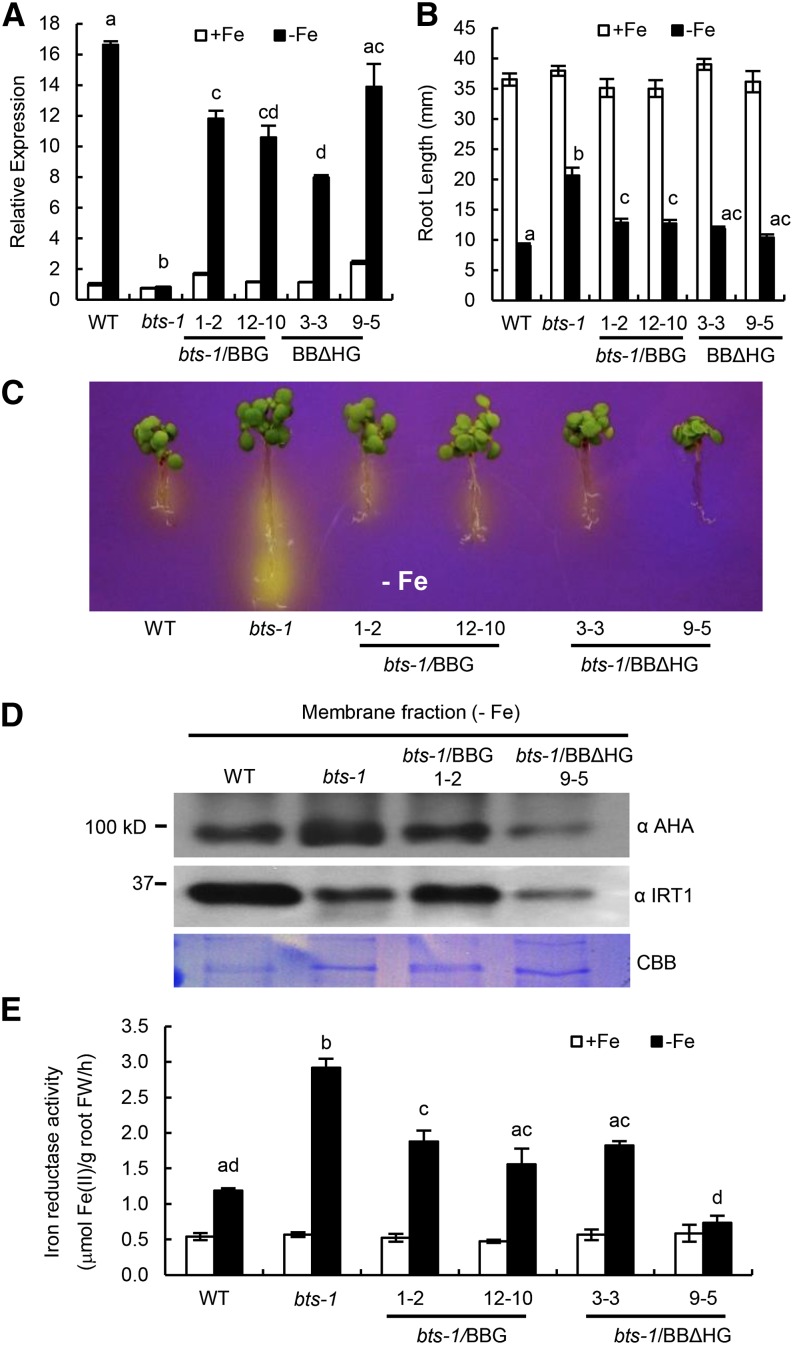
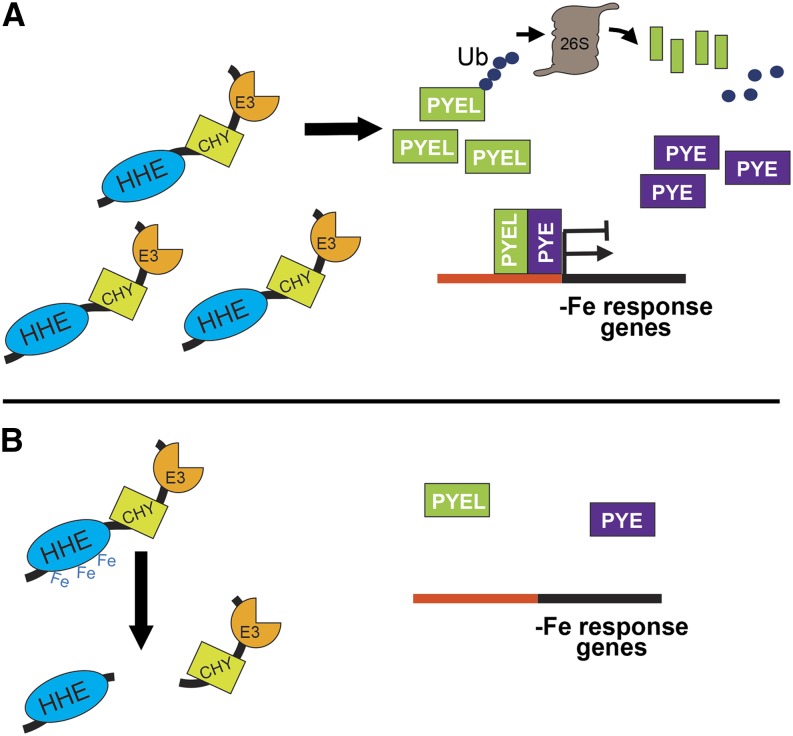
Iron uptake and metabolism are tightly regulated in both plants and animals. In Arabidopsis (Arabidopsis thaliana), BRUTUS (BTS), which contains three hemerythrin (HHE) domains and a Really Interesting New Gene (RING) domain, interacts with basic helix-loop-helix transcription factors that are capable of forming heterodimers with POPEYE (PYE), a positive regulator of the iron deficiency response. BTS has been shown to have E3 ligase capacity and to play a role in root growth, rhizosphere acidification, and iron reductase activity in response to iron deprivation. To further characterize the function of this protein, we examined the expression pattern of recombinant ProBTS::β-GLUCURONIDASE and found that it is expressed in developing embryos and other reproductive tissues, corresponding with its apparent role in reproductive growth and development. Our findings also indicate that the interactions between BTS and PYE-like (PYEL) basic helix-loop-helix transcription factors occur within the nucleus and are dependent on the presence of the RING domain. We provide evidence that BTS facilitates 26S proteasome-mediated degradation of PYEL proteins in the absence of iron. We also determined that, upon binding iron at the HHE domains, BTS is destabilized and that this destabilization relies on specific residues within the HHE domains. This study reveals an important and unique mechanism for plant iron homeostasis whereby an E3 ubiquitin ligase may posttranslationally control components of the transcriptional regulatory network involved in the iron deficiency response.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Berleth T, Jurgens G (1993) The role of the monopteros gene in organising the basal body region of the Arabidopsis embryo. Development 118: 575–587
-
- Besold AN, Lee SJ, Michel SL, Sue NL, Cymet HJ (2010) Functional characterization of iron-substituted neural zinc finger factor 1: metal and DNA binding. J Biol Inorg Chem 15: 583–590 - PubMed
-
- Cabiscol E, Piulats E, Echave P, Herrero E, Ros J (2000) Oxidative stress promotes specific protein damage in Saccharomyces cerevisiae. J Biol Chem 275: 27393–27398 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
