

Dendrites are dispensable for basic motoneuron function but essential for fine tuning of behavior
- PMID: 25453076
- PMCID: PMC4273390
- DOI: 10.1073/pnas.1416247111
Dendrites are dispensable for basic motoneuron function but essential for fine tuning of behavior
Abstract
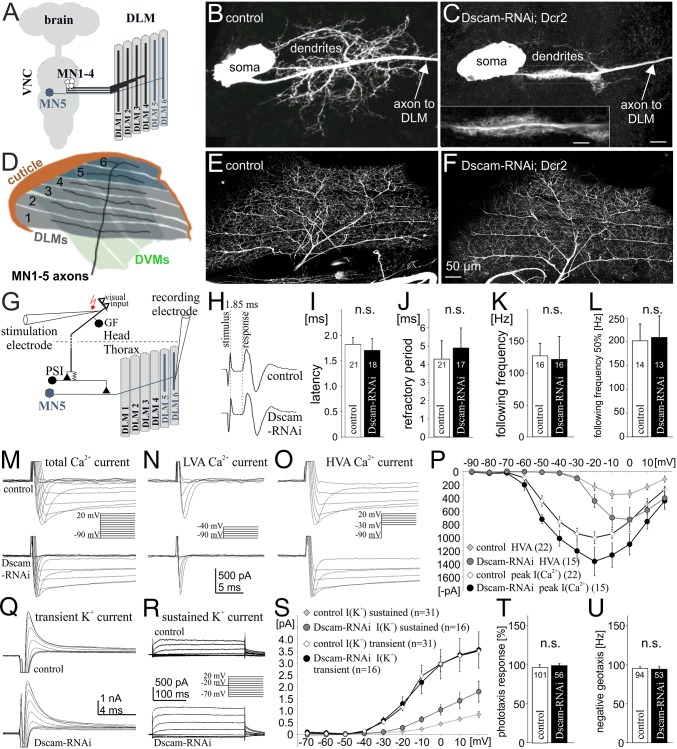
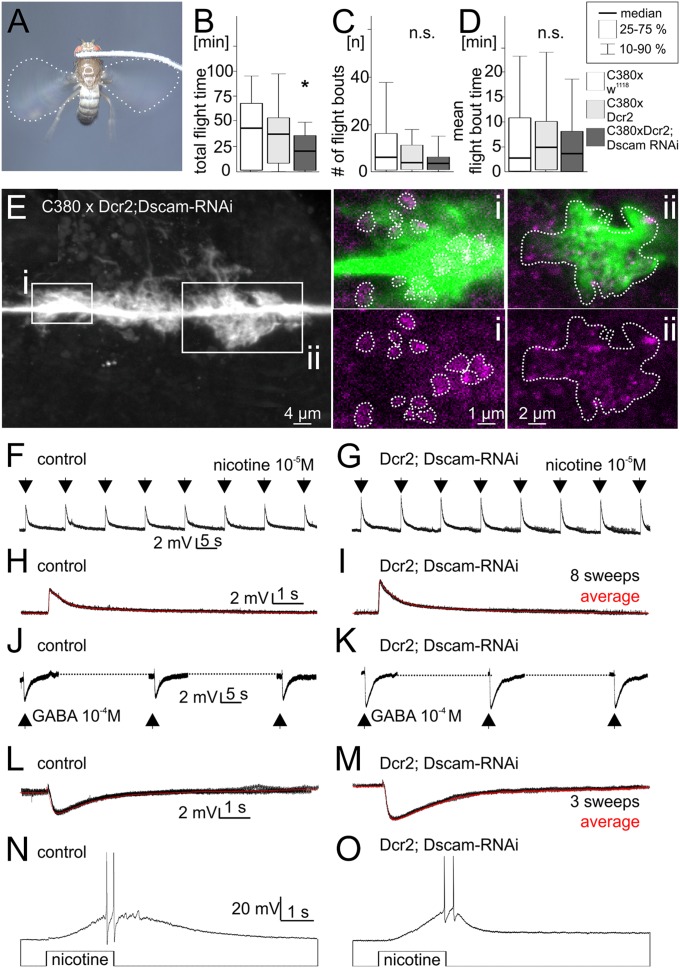
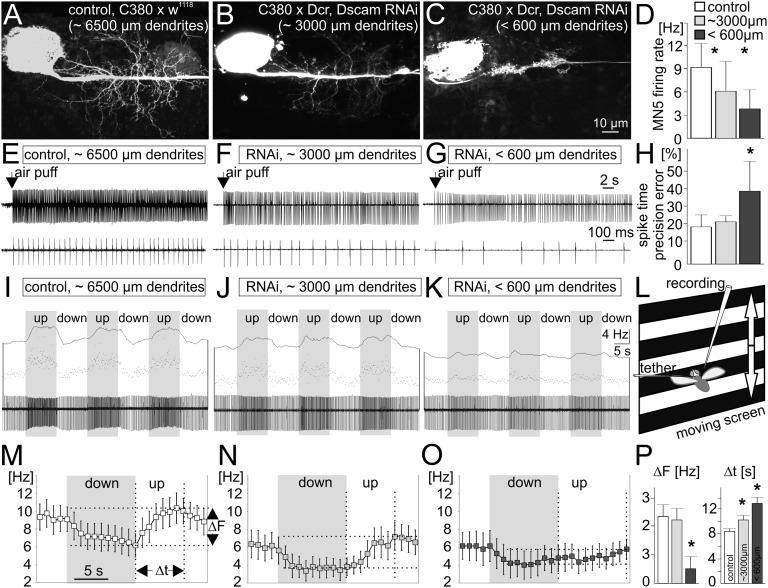
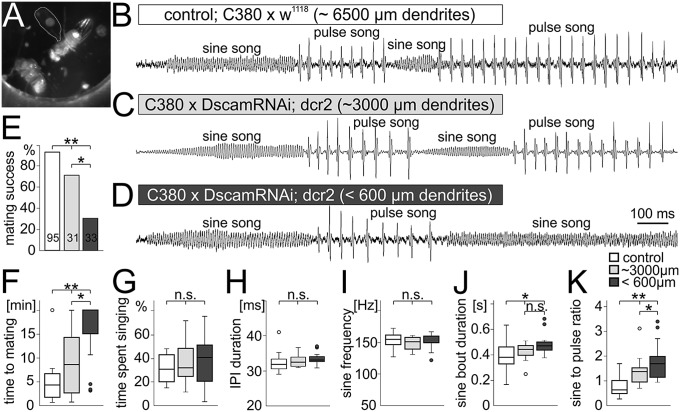
Dendrites are highly complex 3D structures that define neuronal morphology and connectivity and are the predominant sites for synaptic input. Defects in dendritic structure are highly consistent correlates of brain diseases. However, the precise consequences of dendritic structure defects for neuronal function and behavioral performance remain unknown. Here we probe dendritic function by using genetic tools to selectively abolish dendrites in identified Drosophila wing motoneurons without affecting other neuronal properties. We find that these motoneuron dendrites are unexpectedly dispensable for synaptic targeting, qualitatively normal neuronal activity patterns during behavior, and basic behavioral performance. However, significant performance deficits in sophisticated motor behaviors, such as flight altitude control and switching between discrete courtship song elements, scale with the degree of dendritic defect. To our knowledge, our observations provide the first direct evidence that complex dendrite architecture is critically required for fine-tuning and adaptability within robust, evolutionarily constrained behavioral programs that are vital for mating success and survival. We speculate that the observed scaling of performance deficits with the degree of structural defect is consistent with gradual increases in intellectual disability during continuously advancing structural deficiencies in progressive neurological disorders.
Keywords: Drosophila; courtship; dendrite; motor behavior; synapse.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Fiala JC, Spacek J, Harris KM. Dendritic structure. In: Stuart G, Sprouston N, Häusser M, editors. Dendrites. Oxford University Press; New York: 2008. pp. 1–34.
-
- Azevedo FA, et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J Comp Neurol. 2009;513(5):532–541. - PubMed
-
- Gabbiani F, Krapp HG, Koch C, Laurent G. Multiplicative computation in a visual neuron sensitive to looming. Nature. 2002;420(6913):320–324. - PubMed
-
- Single S, Borst A. Dendritic integration and its role in computing image velocity. Science. 1998;281(5384):1848–1850. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
