

The yeast polo kinase Cdc5 regulates the shape of the mitotic nucleus
- PMID: 25454593
- PMCID: PMC4255140
- DOI: 10.1016/j.cub.2014.10.029
The yeast polo kinase Cdc5 regulates the shape of the mitotic nucleus
Abstract
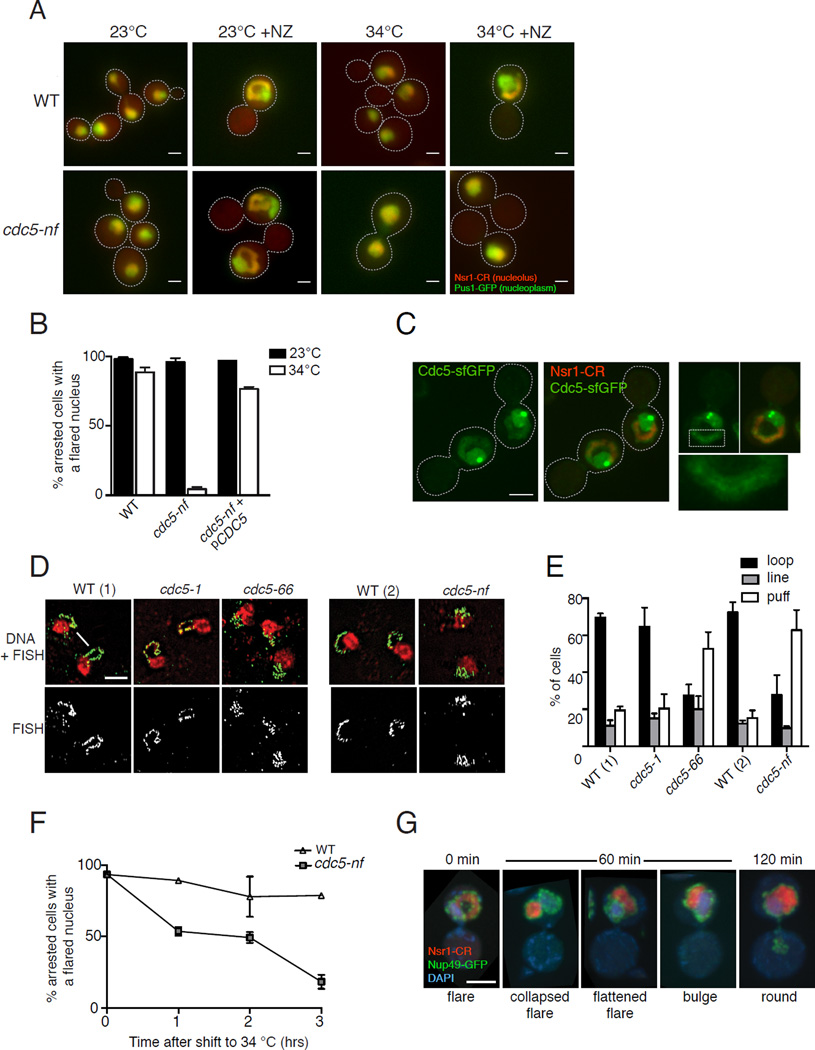
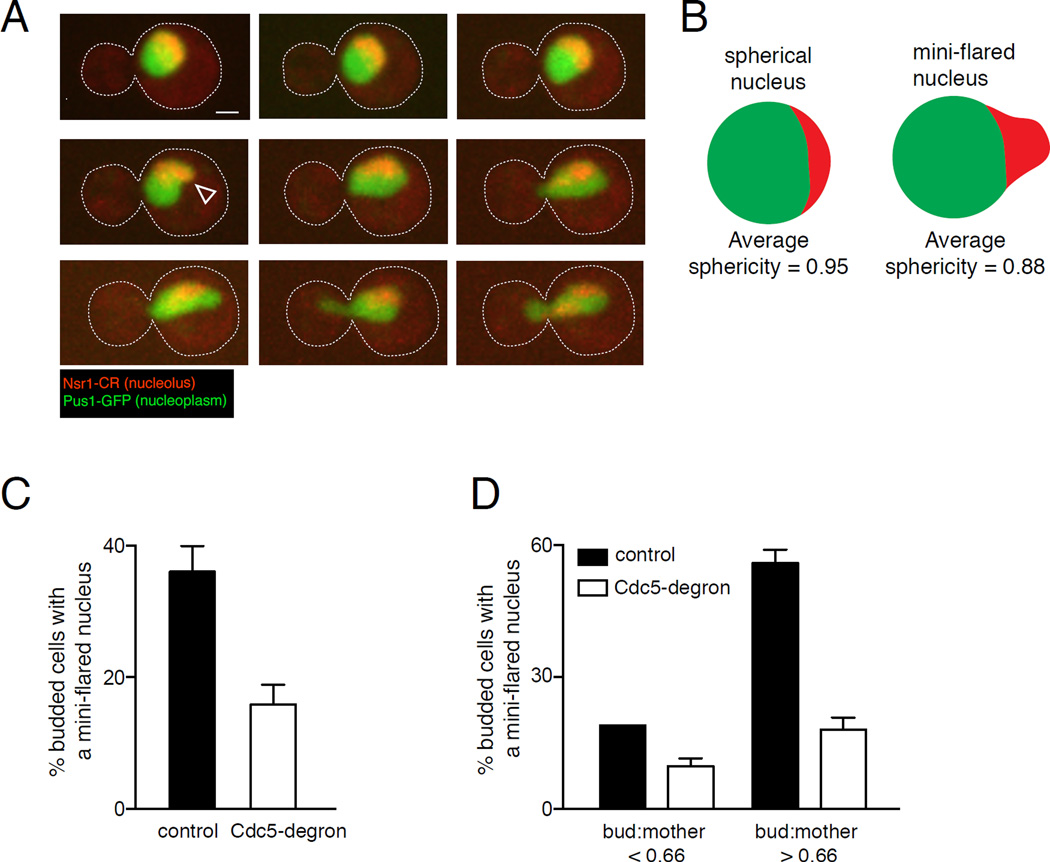
Abnormal nuclear size and shape are hallmarks of aging and cancer. However, the mechanisms regulating nuclear morphology and nuclear envelope (NE) expansion are poorly understood. In metazoans, the NE disassembles prior to chromosome segregation and reassembles at the end of mitosis. In budding yeast, the NE remains intact. The nucleus elongates as chromosomes segregate and then divides at the end of mitosis to form two daughter nuclei without NE disassembly. The budding yeast nucleus also undergoes remodeling during a mitotic arrest; the NE continues to expand despite the pause in chromosome segregation, forming a nuclear extension, or "flare," that encompasses the nucleolus. The distinct nucleolar localization of the mitotic flare indicates that the NE is compartmentalized and that there is a mechanism by which NE expansion is confined to the region adjacent to the nucleolus. Here we show that mitotic flare formation is dependent on the yeast polo kinase Cdc5. This function of Cdc5 is independent of its known mitotic roles, including rDNA condensation. High-resolution imaging revealed that following Cdc5 inactivation, nuclei expand isometrically rather than forming a flare, indicating that Cdc5 is needed for NE compartmentalization. Even in an uninterrupted cell cycle, a small NE expansion occurs adjacent to the nucleolus prior to anaphase in a Cdc5-dependent manner. Our data provide the first evidence that polo kinase, a key regulator of mitosis, plays a role in regulating nuclear morphology and NE expansion.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures


References
-
- Archambault V, Glover DM. Polo-like kinases: conservation and divergence in their functions and regulation. Nat Rev Mol Cell Biol. 2009;10:265–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
