

RanBP1 governs spindle assembly by defining mitotic Ran-GTP production
- PMID: 25458009
- PMCID: PMC4254504
- DOI: 10.1016/j.devcel.2014.10.014
RanBP1 governs spindle assembly by defining mitotic Ran-GTP production
Abstract
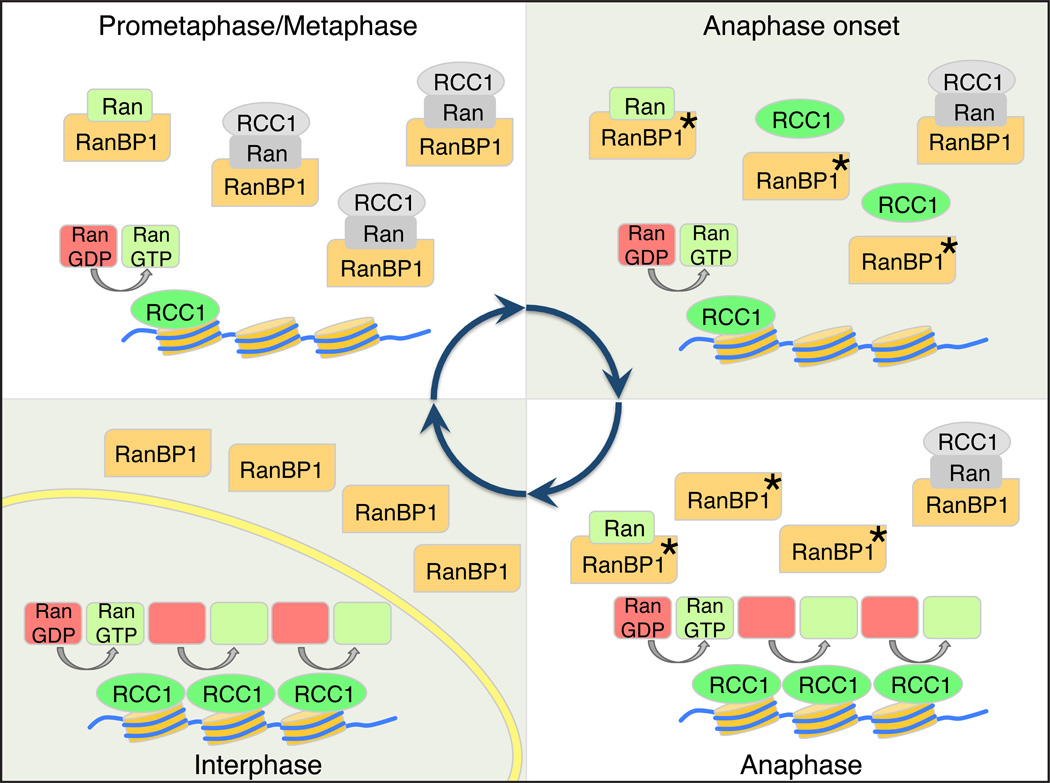
Accurate control of the Ras-related nuclear protein (Ran) GTPase cycle depends on the regulated activity of regulator of chromosome condensation 1 (RCC1), Ran's nucleotide exchange factor. RanBP1 has been characterized as a coactivator of the Ran GTPase-activating protein RanGAP1. RanBP1 can also form a stable complex with Ran and RCC1, although the dynamics and function of this complex remain poorly understood. Here, we show that formation of the heterotrimeric RCC1/Ran/RanBP1 complex in M phase Xenopus egg extracts controls both RCC1's enzymatic activity and partitioning between the chromatin-bound and soluble pools of RCC1. This mechanism is critical for spatial control of Ran-guanosine triphosphate (GTP) gradients that guide mitotic spindle assembly. Moreover, phosphorylation of RanBP1 drives changes in the dynamics of chromatin-bound RCC1 pools at the metaphase-anaphase transition. Our findings reveal an important mitotic role for RanBP1, controlling the spatial distribution and magnitude of mitotic Ran-GTP production and thereby ensuring accurate execution of Ran-dependent mitotic events.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
RanBP1 controls the Ran pathway in mammalian cells through regulation of mitotic RCC1 dynamics.Cell Cycle. 2020 Aug;19(15):1899-1916. doi: 10.1080/15384101.2020.1782036. Epub 2020 Jun 28. Cell Cycle. 2020. PMID: 32594833 Free PMC article.
-
The ran GTPase regulates mitotic spindle assembly.Curr Biol. 1999 May 6;9(9):481-4. doi: 10.1016/s0960-9822(99)80213-9. Curr Biol. 1999. PMID: 10322113
-
The Ran GTPase regulates kinetochore function.Dev Cell. 2003 Jul;5(1):99-111. doi: 10.1016/s1534-5807(03)00194-1. Dev Cell. 2003. PMID: 12852855
-
Ran-GTP regulates kinetochore attachment in somatic cells.Cell Cycle. 2005 Sep;4(9):1161-5. doi: 10.4161/cc.4.9.1979. Epub 2005 Sep 28. Cell Cycle. 2005. PMID: 16082212 Review.
-
A new role of ran GTPase.Biochem Biophys Res Commun. 1999 Sep 7;262(3):571-4. doi: 10.1006/bbrc.1999.1252. Biochem Biophys Res Commun. 1999. PMID: 10471364 Review.
Cited by
-
Nup50 plays more than one instrument.Cell Cycle. 2022 Sep;21(17):1785-1794. doi: 10.1080/15384101.2022.2074742. Epub 2022 May 13. Cell Cycle. 2022. PMID: 35549614 Free PMC article.
-
Studying the variations in differently expressed serum proteins of Hainan black goat during the breeding cycle using isobaric tags for relative and absolute quantitation (iTRAQ) technology.J Reprod Dev. 2019 Oct 23;65(5):413-421. doi: 10.1262/jrd.2018-105. Epub 2019 Jul 14. J Reprod Dev. 2019. PMID: 31308307 Free PMC article.
-
Unveiling the Movement of RanBP1 During the Cell Cycle and Its Interaction with a Cyclin-Dependent Kinase (CDK) in Plants.Int J Mol Sci. 2024 Dec 24;26(1):46. doi: 10.3390/ijms26010046. Int J Mol Sci. 2024. PMID: 39795905 Free PMC article.
-
Human ectodermal organoids reveal the cellular origin of DiGeorge Syndrome.bioRxiv [Preprint]. 2025 Aug 8:2025.08.08.669417. doi: 10.1101/2025.08.08.669417. bioRxiv. 2025. PMID: 40855884 Free PMC article. Preprint.
-
Spatiotemporal 22q11.21 Protein Network Implicates DGCR8-Dependent MicroRNA Biogenesis as a Risk for Late-Fetal Cortical Development in Psychiatric Diseases.Life (Basel). 2021 May 31;11(6):514. doi: 10.3390/life11060514. Life (Basel). 2021. PMID: 34073122 Free PMC article.
References
-
- Amato R, Scumaci D, D'Antona L, Iuliano R, Menniti M, Di Sanzo M, Faniello MC, Colao E, Malatesta P, Zingone A, et al. Sgk1 enhances RANBP1 transcript levels and decreases taxol sensitivity in RKO colon carcinoma cells. Oncogene. 2013;32:4572–4578. - PubMed
-
- Arnaoutov A, Dasso M. The Ran GTPase regulates kinetochore function. Developmental cell. 2003;5:99–111. - PubMed
-
- Bischoff FR, Gorlich D. RanBP1 is crucial for the release of RanGTP from importin beta-related nuclear transport factors. FEBS letters. 1997;419:249–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
