

Deciphering the evolutionary history of open and closed mitosis
- PMID: 25458223
- PMCID: PMC4278198
- DOI: 10.1016/j.cub.2014.10.011
Deciphering the evolutionary history of open and closed mitosis
Abstract
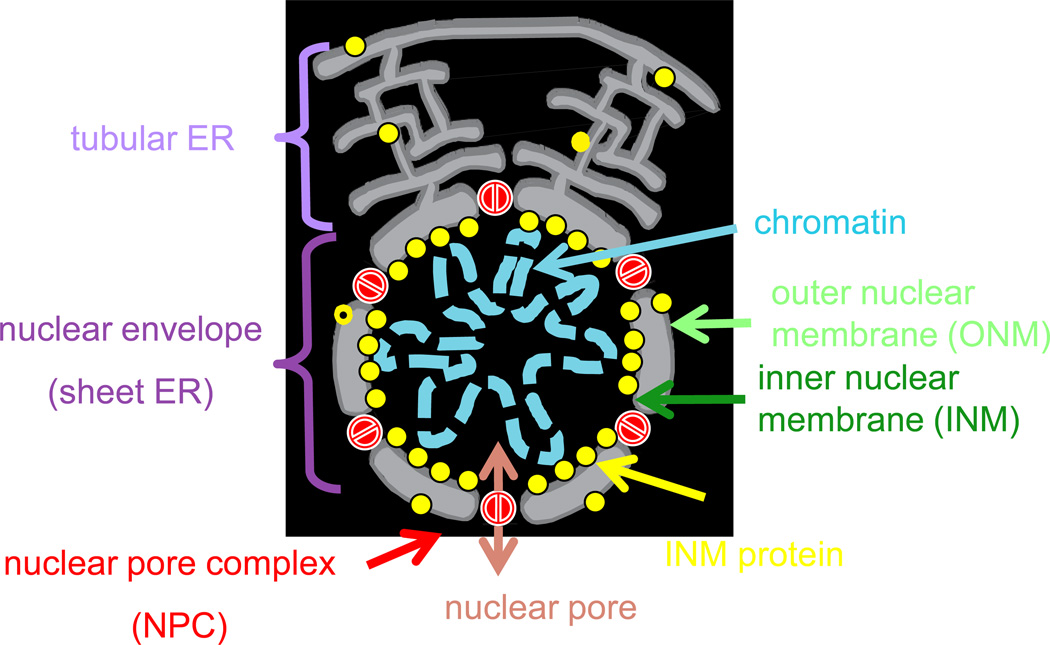
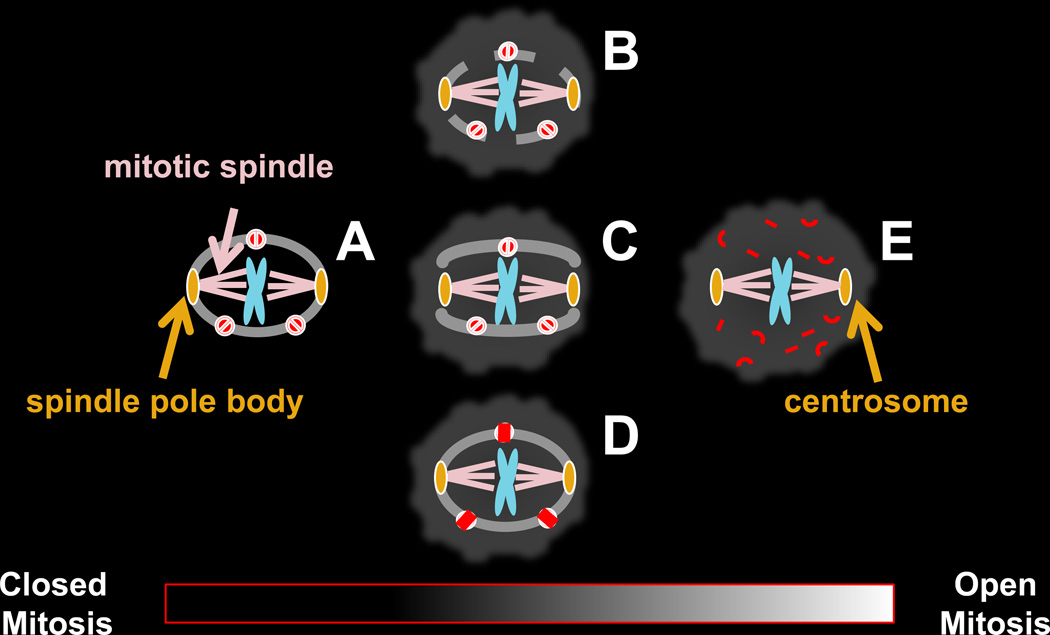
The origin of the nucleus at the prokaryote-to-eukaryote transition represents one of the most important events in the evolution of cellular organization. The nuclear envelope encircles the chromosomes in interphase and is a selectively permeable barrier between the nucleoplasm and cytoplasm and an organizational scaffold for the nucleus. It remains intact in the 'closed' mitosis of some yeasts, but loses its integrity in the 'open' mitosis of mammals. Instances of both types of mitosis within two evolutionary clades indicate multiple evolutionary transitions between open and closed mitosis, although the underlying genetic changes that influenced these transitions remain unknown. A survey of the diversity of mitotic nuclei that fall between these extremes is the starting point from which to determine the physiologically relevant characteristics distinguishing open from closed mitosis and to understand how they evolved and why they are retained in present-day organisms. The field is now poised to begin addressing these issues by defining and documenting patterns of mitotic nuclear variation within and among species and mapping them onto a phylogenic tree. Deciphering the evolutionary history of open and closed mitosis will complement cell biological and genetic approaches aimed at deciphering the fundamental organizational principles of the nucleus.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Allwood AC, Walter MR, Kamber BS, Marshall CP, Burch IW. Stromatolite reef from the Early Archaean era of Australia. Nature. 2006;441:714–718. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
