

Olfactory system oscillations across phyla
- PMID: 25460070
- PMCID: PMC4374988
- DOI: 10.1016/j.conb.2014.10.004
Olfactory system oscillations across phyla
Abstract
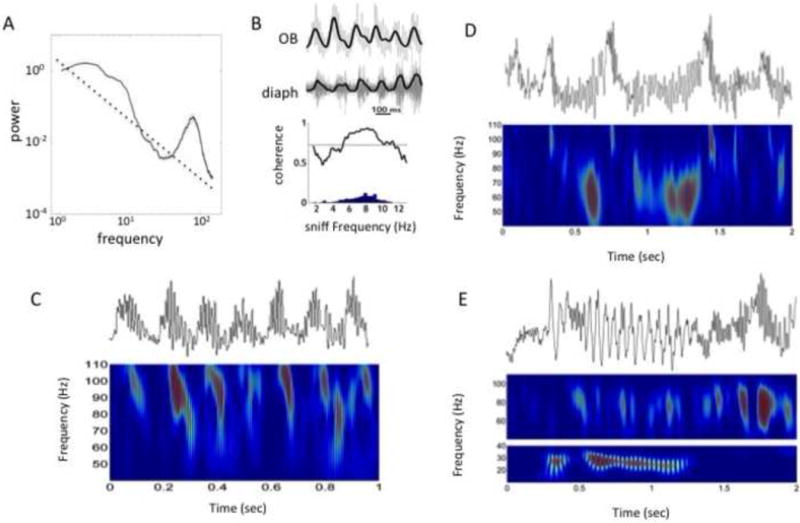
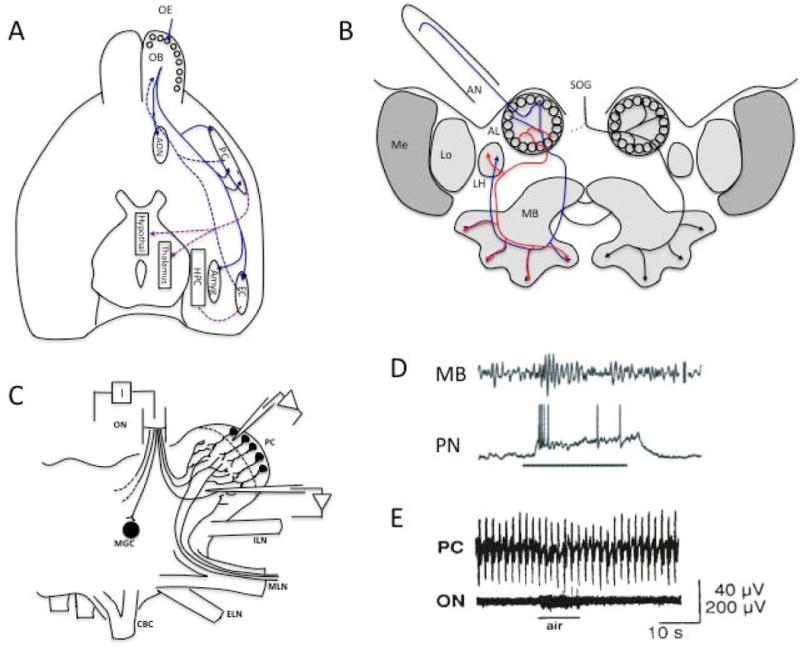
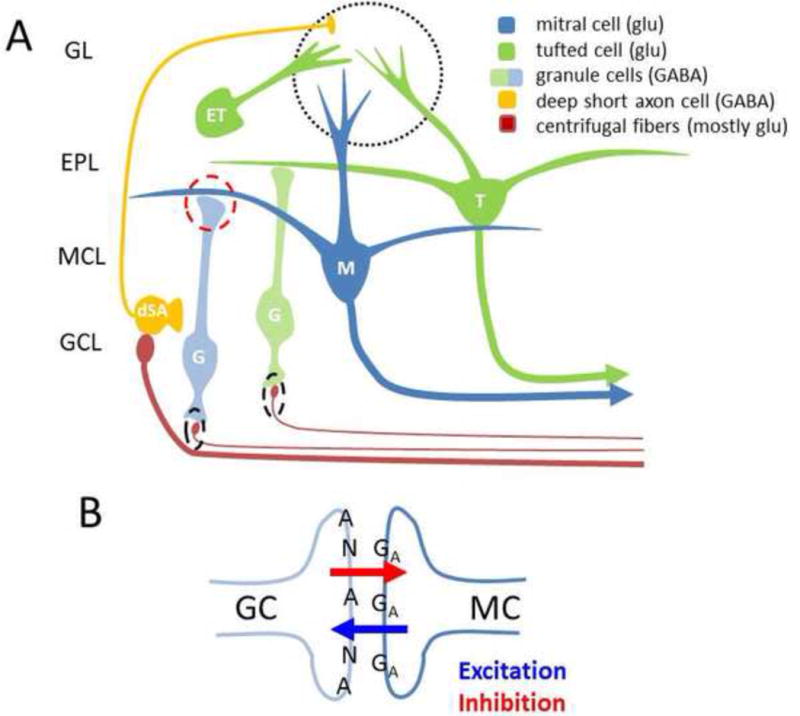
Neural oscillations are ubiquitous in olfactory systems of mammals, insects and molluscs. Neurophysiological and computational investigations point to common mechanisms for gamma or odor associated oscillations across phyla (40-100Hz in mammals, 20-30Hz in insects, 0.5-1.5Hz in molluscs), engaging the reciprocal dendrodendritic synapse between excitatory principle neurons and inhibitory interneurons in the olfactory bulb (OB), antennal lobe (AL), or procerebrum (PrC). Recent studies suggest important mechanisms that may modulate gamma oscillations, including neuromodulators and centrifugal input to the OB and AL. Beta (20Hz) and theta (2-12Hz) oscillations coordinate activity within and across brain regions. Olfactory beta oscillations are associated with odor learning and depend on centrifugal OB input, while theta oscillations are strongly associated with respiration.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of Interest Statement
Nothing to declare
Figures
Similar articles
-
Circuit oscillations in odor perception and memory.Prog Brain Res. 2014;208:223-51. doi: 10.1016/B978-0-444-63350-7.00009-7. Prog Brain Res. 2014. PMID: 24767485 Review.
-
Localization of odor-induced oscillations in the bumblebee antennal lobe.Neurosci Lett. 2001 Dec 28;316(3):133-6. doi: 10.1016/s0304-3940(01)02385-0. Neurosci Lett. 2001. PMID: 11744220
-
Olfactory bulb gamma oscillations are enhanced with task demands.J Neurosci. 2007 Aug 1;27(31):8358-65. doi: 10.1523/JNEUROSCI.1199-07.2007. J Neurosci. 2007. PMID: 17670982 Free PMC article.
-
In vivo recordings of spontaneous and odor-modulated dynamics in the Limax olfactory lobe.J Neurobiol. 2001 Feb 5;46(2):126-41. J Neurobiol. 2001. PMID: 11153014
-
Newborn neurons in the adult olfactory bulb: unique properties for specific odor behavior.Behav Brain Res. 2012 Feb 14;227(2):480-9. doi: 10.1016/j.bbr.2011.08.001. Epub 2011 Aug 6. Behav Brain Res. 2012. PMID: 21843557 Review.
Cited by
-
Snapshots of the Brain in Action: Local Circuit Operations through the Lens of γ Oscillations.J Neurosci. 2016 Oct 12;36(41):10496-10504. doi: 10.1523/JNEUROSCI.1021-16.2016. J Neurosci. 2016. PMID: 27733601 Free PMC article.
-
Network mechanism for insect olfaction.Cogn Neurodyn. 2021 Feb;15(1):103-129. doi: 10.1007/s11571-020-09640-3. Epub 2021 Jan 15. Cogn Neurodyn. 2021. PMID: 33786083 Free PMC article.
-
Breathing as a Fundamental Rhythm of Brain Function.Front Neural Circuits. 2017 Jan 12;10:115. doi: 10.3389/fncir.2016.00115. eCollection 2016. Front Neural Circuits. 2017. PMID: 28127277 Free PMC article. Review.
-
GABAergic modulation of olfactomotor transmission in lampreys.PLoS Biol. 2018 Oct 4;16(10):e2005512. doi: 10.1371/journal.pbio.2005512. eCollection 2018 Oct. PLoS Biol. 2018. PMID: 30286079 Free PMC article.
-
Excitability of Neural Activity is Enhanced, but Neural Discrimination of Odors is Slightly Decreased, in the Olfactory Bulb of Fasted Mice.Genes (Basel). 2020 Apr 16;11(4):433. doi: 10.3390/genes11040433. Genes (Basel). 2020. PMID: 32316323 Free PMC article.
References
-
- Eisthen HL. Why are olfactory systems of different animals so similar? [Internet] Brain Behav Evol. 2002;59:273–293. - PubMed
-
- Adrian ED. The electrical activity of the mammalian olfactory bulb. EEG Clin Neurophysiol. 1950;2:377–388. - PubMed
-
- Freeman WJ. Linear distributed feedback model for prepyriform cortex. Exp Neurol. 1964;10:525–547. - PubMed
-
- Rall W, Shepherd GM. Theoretical reconstruction dendrodendritic of rield potentials and in olfactory bulb synaptic interactions. J Neurophysiol. 1968;31:884–915. - PubMed
-
- Bressler SL, Freeman WJ. Frequency analysis of olfactory system EEG in cat, rabbit, and rat [Internet] Electroencephalogr Clin Neurophysiol. 1980;50:19–24. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
