

Review
doi: 10.1016/j.sbi.2014.09.006.
Epub 2014 Oct 14.
Structure and mechanism of lanthipeptide biosynthetic enzymes
Affiliations
- PMID: 25460269
- PMCID: PMC4267917
- DOI: 10.1016/j.sbi.2014.09.006
Item in Clipboard
Review
Structure and mechanism of lanthipeptide biosynthetic enzymes
Curr Opin Struct Biol.
2014 Dec.
Abstract
Lanthipeptides are members of the ribosomally synthesized and post-translationally modified peptide (RiPP) natural products. They contain thioether crosslinks generated by dehydration of Ser and Thr residues followed by the addition of the thiol of Cys residues to the dehydroamino acids. Recent studies have revealed unexpected mechanisms of the post-translational modifications, and structural studies have started to provide insights into recognition of the peptide substrates by the modification enzymes.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
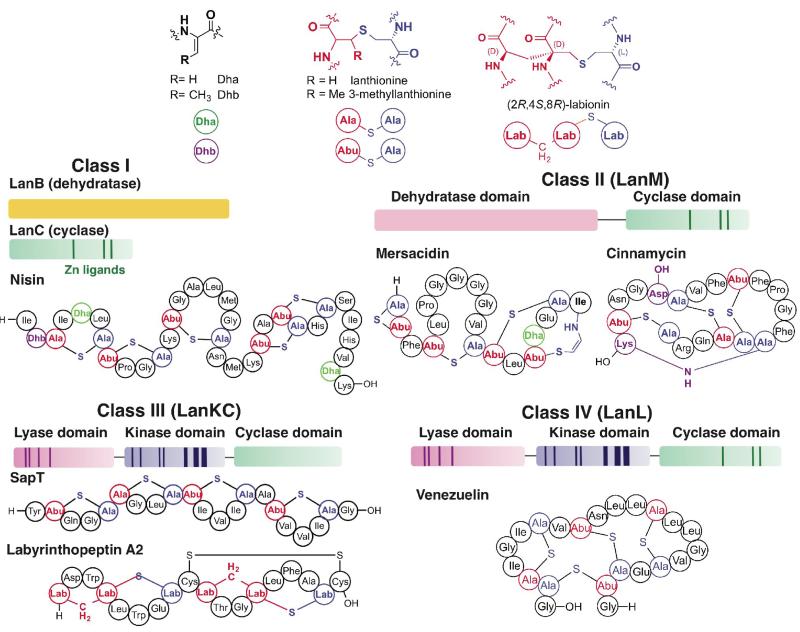
Structures of the canonical post-translational modifications in lanthipeptides, overview of the four classes of lanthipeptide biosynthesis, and structures of representative members. The class I lantibiotics, such as nisin, are generated by two independent enzymes (LanB dehydratase and LanC cyclase); class II molecules like mersacidin and cinnamycin are biosynthesized by a bifunctional enzyme that contains both dehydration and cyclization activities; the biosynthesis of class III molecules, such as SapT or labyrinthopeptin A2, is carried out by a single protein that has lyase, kinase, and cyclase domains but lacks the signature zinc-ligands in the cyclase domain. Some class III lanthipeptides contain labionin moieties; class IV lantibiotics, such as venezuelin are generated by an enzyme that also contains lyase and kinase domains but a LanC-like cyclase domain that contains the metal ligands.
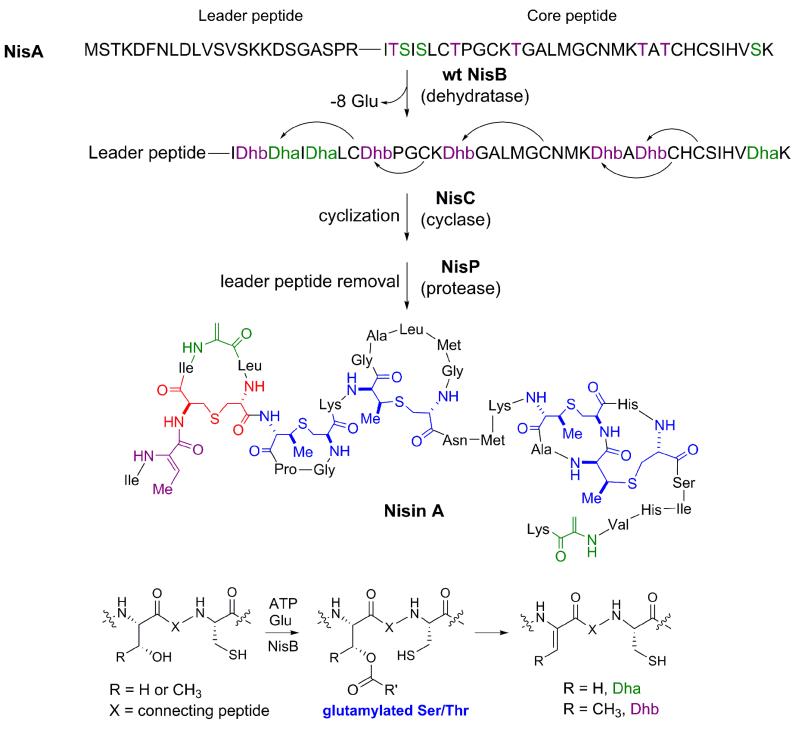
The biosynthesis of the class I lantibiotic nisin is catalyzed by a dehydratase NisB and a cyclase NisC. The activation of Ser/Thr by the addition of glutamate was uncovered by analysis of NisB mutants. Wild type NisB is able to eliminate the glutamates to generate the dehydroamino acid residues. During normal catalysis, the glutamates are cryptic and never observed. The NisC cyclase carries out cyclization in a zinc-dependent manner but the molecular mechanism that governs site specificity for thioether formation is not yet known. The leader peptide is removed from the modified peptide by a specific protease to yield the bioactive natural product. Lanthionines are shown in red, methyllanthionines in blue.
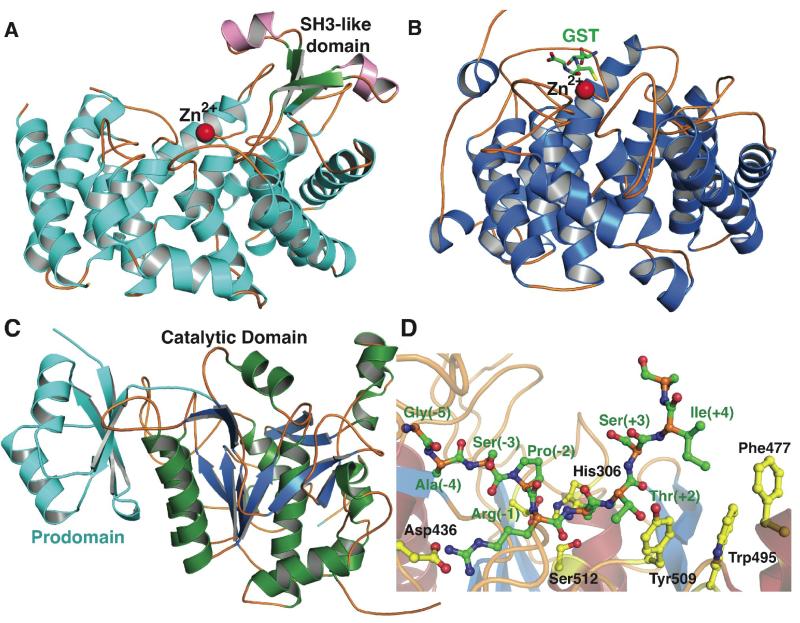
Structures of the nisin cyclase NisC, the eukaryotic lanthionine synthase C-like protein LanCL1, a lanthipeptide protease-like protein from S. aureus, and the protease NisP. (A) NisC contains a structural element resembling eukaryotic peptide binding domains that is missing in the LanCL proteins such as LanCL1 (B). (C) Crystal structure of the orphan lantibiotic protease homolog from S. aureus with the prodomain colored in cyan and the catalytic domain colored in green and blue. (D) A model of the NisP protease in complex with an unmodified NisA peptide substrate, generated using the S. aureus orphan enzyme as a template. Protease residues are shown in yellow and substrate residues are shown in green. The presence of an aromatic wall that blocks the carboxy- terminal end of the substrate-binding site may facilitate a preference for modified substrates containing a thioether cycle.
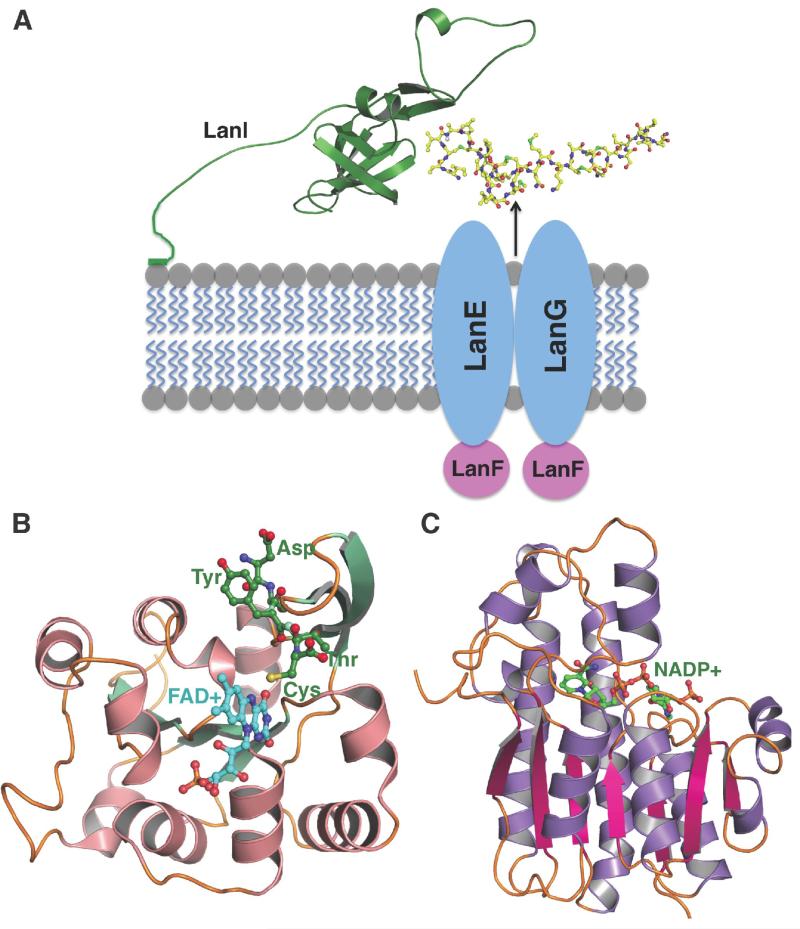
Structures of lanthipeptide immunity proteins and tailoring enzymes. (A) The coordinated actions of the LanFEG transport system and the LanI lantibiotic-binding protein confer self-immunity against lantibiotic toxicity in producing strains. The depicted orientation of the immunity protein is based upon the NMR structure of SpaI and biochemical data that suggest that the amino-terminus of the immunity protein binds to lipid membranes even in the absence of a covalent anchor. Presumably, the immunity protein would act to bind directly to lantibiotics that are then expelled from the cell through the FEG transport system. While several lanthipeptide tailoring enzymes have been characterized, structural data are available only for (B) oxidative decarboxylases, illustrated for EpiD (shown in pink and green) involved in the maturation of epidermin, and (C) the short-chain dehydrogenase ElxO that processes epilancin 15X (shown in purple and red).
References
-
- Clardy J, Fischbach MA, Walsh CT. New antibiotics from bacterial natural products. Nat Biotechnol. 2006;24:1541–1550. - PubMed
-
- Arnison PG, Bibb MJ, Bierbaum G, Bowers AA, Bugni TS, Bulaj G, Camarero JA, Campopiano DJ, Challis GL, Clardy J, et al. Ribosomally Synthesized and Post-translationally Modified Peptide Natural Products: Overview and Recommendations for a Universal Nomenclature. Nat. Prod. Rep. 2013;30:108–160. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
