

Metformin protects skeletal muscle from cardiotoxin induced degeneration
- PMID: 25461598
- PMCID: PMC4252070
- DOI: 10.1371/journal.pone.0114018
Metformin protects skeletal muscle from cardiotoxin induced degeneration
Abstract
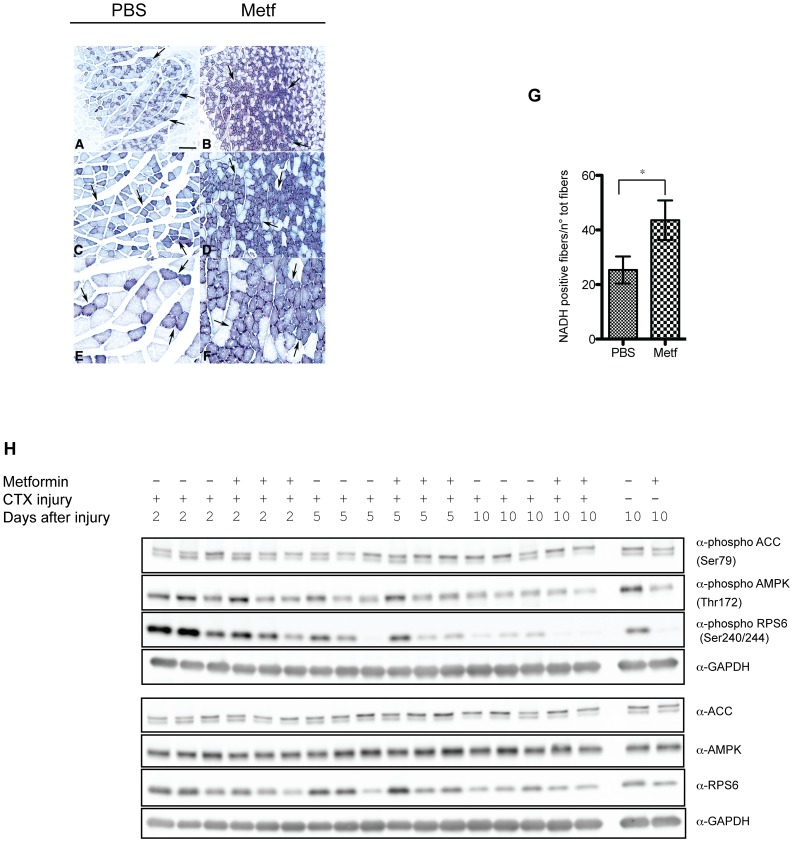
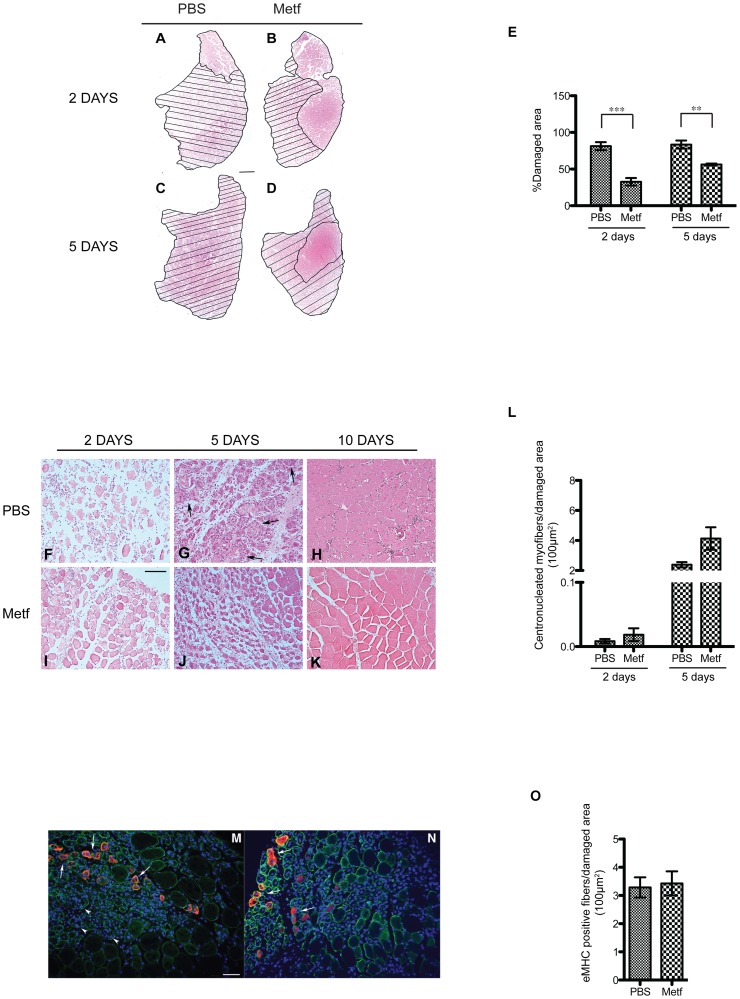
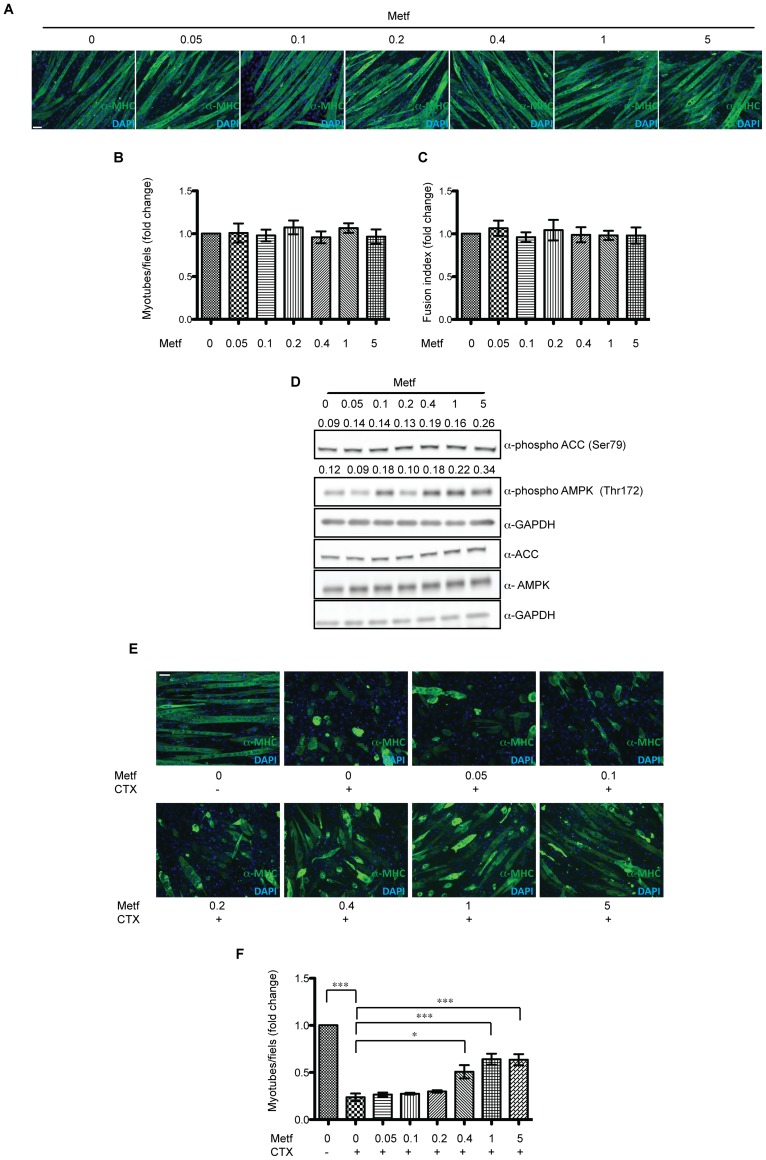
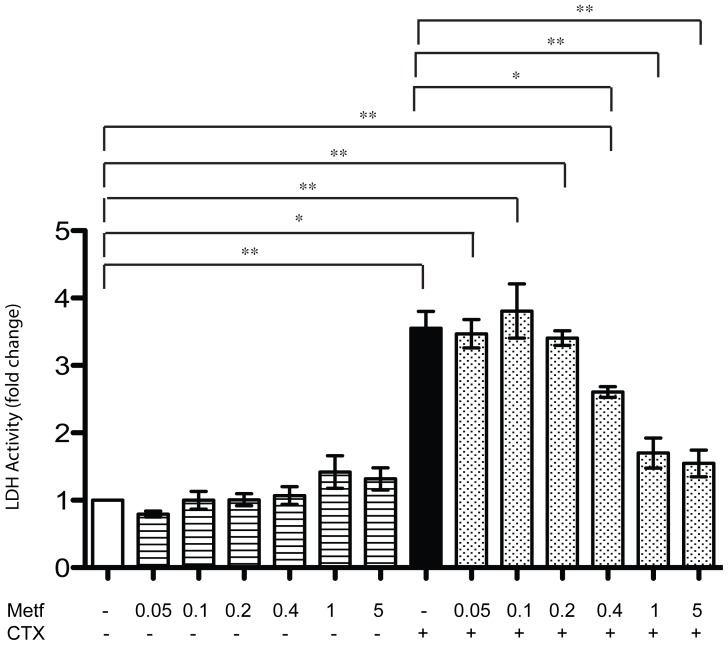
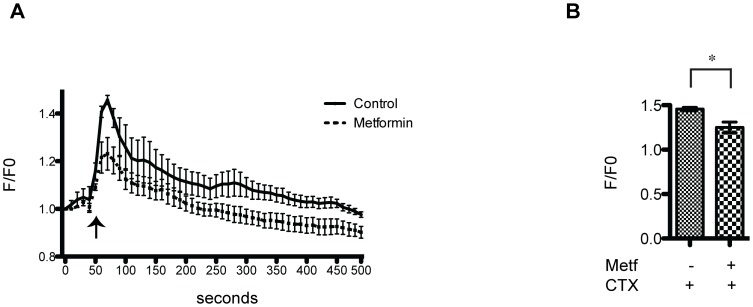
The skeletal muscle tissue has a remarkable capacity to regenerate upon injury. Recent studies have suggested that this regenerative process is improved when AMPK is activated. In the muscle of young and old mice a low calorie diet, which activates AMPK, markedly enhances muscle regeneration. Remarkably, intraperitoneal injection of AICAR, an AMPK agonist, improves the structural integrity of muscles of dystrophin-deficient mdx mice. Building on these observations we asked whether metformin, a powerful anti-hyperglycemic drug, which indirectly activates AMPK, affects the response of skeletal muscle to damage. In our conditions, metformin treatment did not significantly influence muscle regeneration. On the other hand we observed that the muscles of metformin treated mice are more resilient to cardiotoxin injury displaying lesser muscle damage. Accordingly myotubes, originated in vitro from differentiated C2C12 myoblast cell line, become more resistant to cardiotoxin damage after pre-incubation with metformin. Our results indicate that metformin limits cardiotoxin damage by protecting myotubes from necrosis. Although the details of the molecular mechanisms underlying the protective effect remain to be elucidated, we report a correlation between the ability of metformin to promote resistance to damage and its capacity to counteract the increment of intracellular calcium levels induced by cardiotoxin treatment. Since increased cytoplasmic calcium concentrations characterize additional muscle pathological conditions, including dystrophies, metformin treatment could prove a valuable strategy to ameliorate the conditions of patients affected by dystrophies.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
