

Homeostasis of phospholipids - The level of phosphatidylethanolamine tightly adapts to changes in ethanolamine plasmalogens
- PMID: 25463479
- PMCID: PMC4331674
- DOI: 10.1016/j.bbalip.2014.11.005
Homeostasis of phospholipids - The level of phosphatidylethanolamine tightly adapts to changes in ethanolamine plasmalogens
Abstract
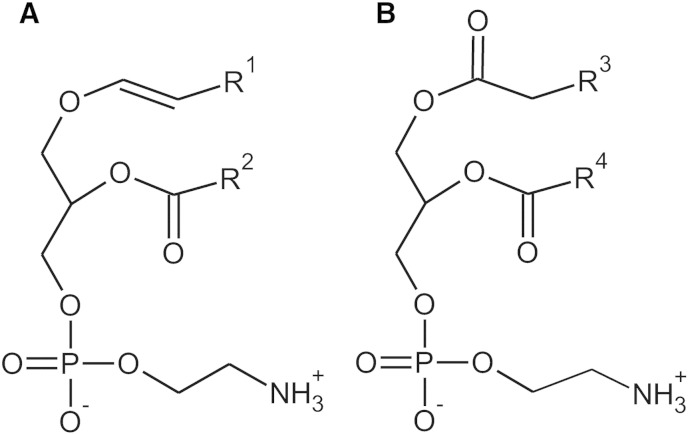
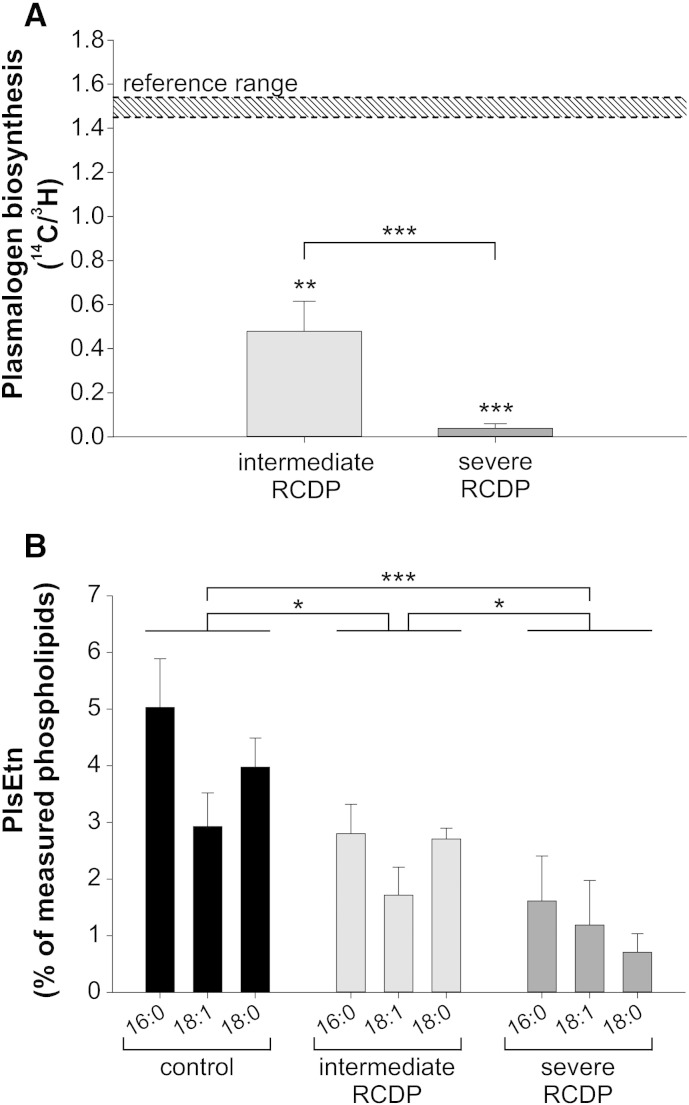
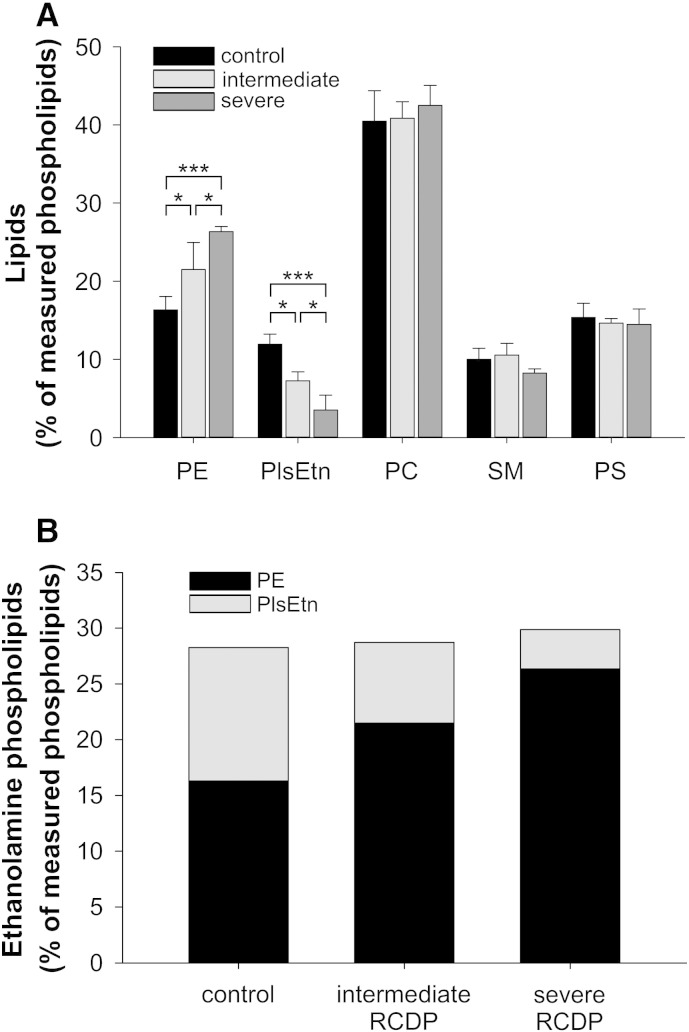
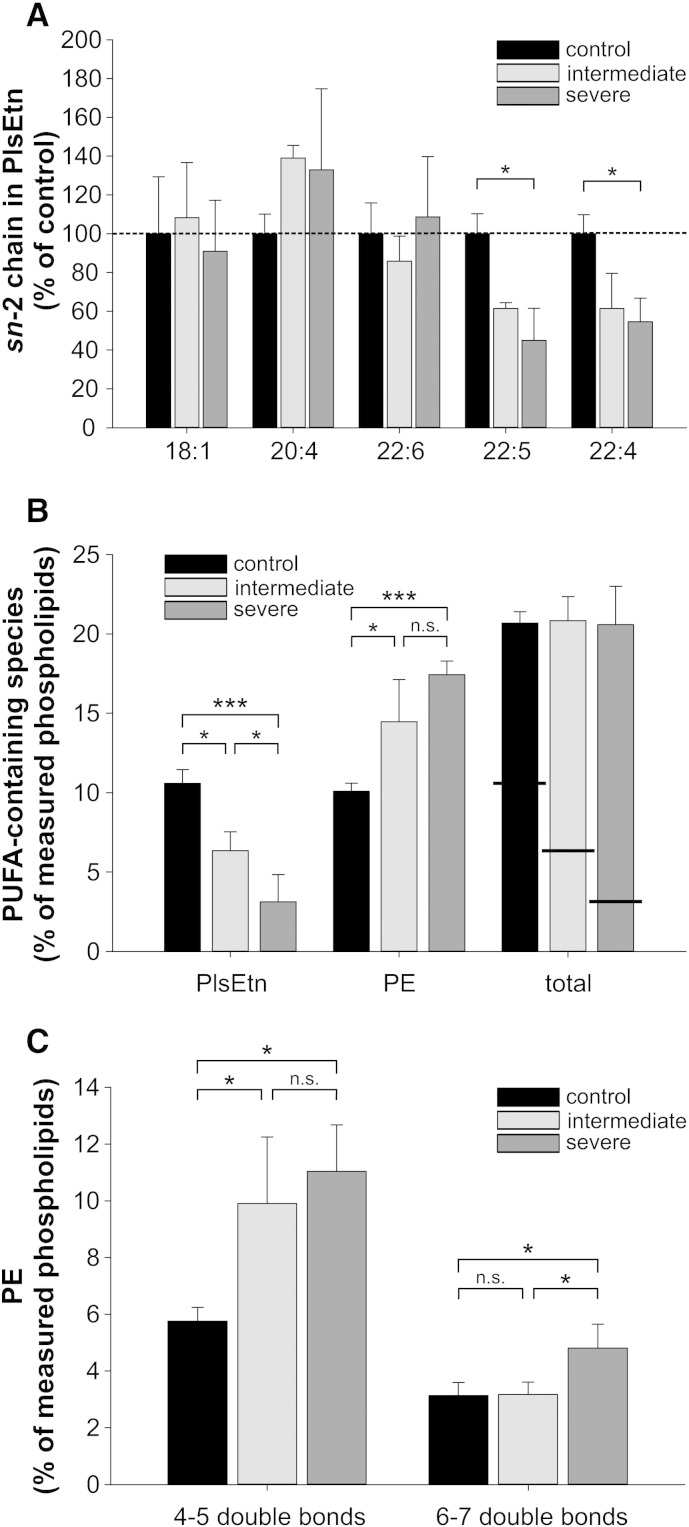
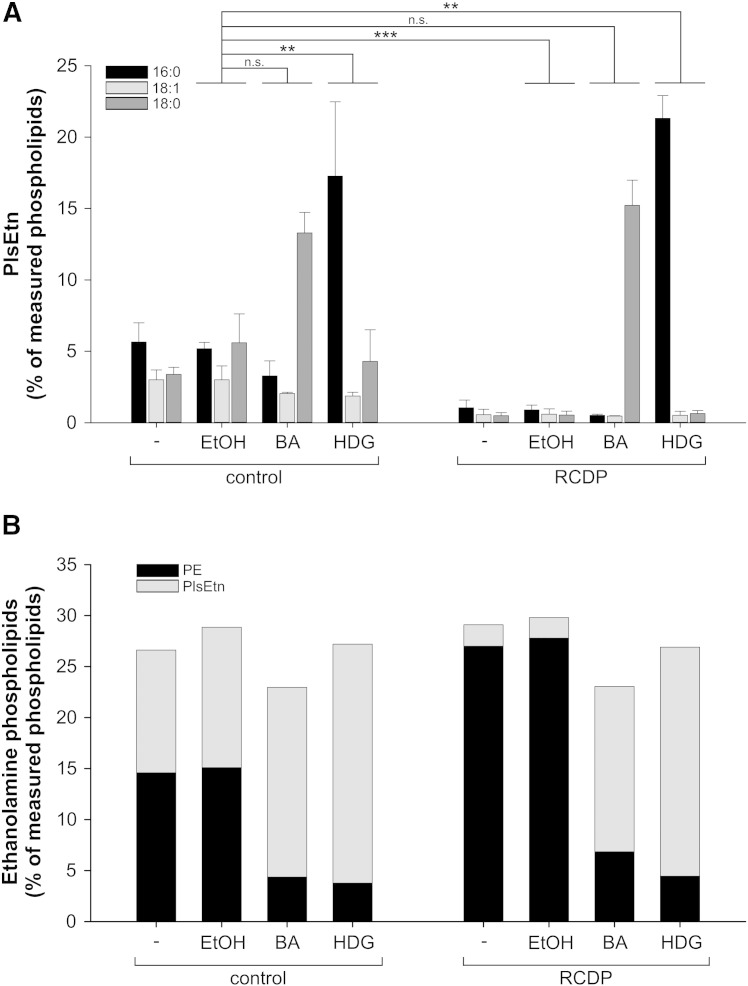
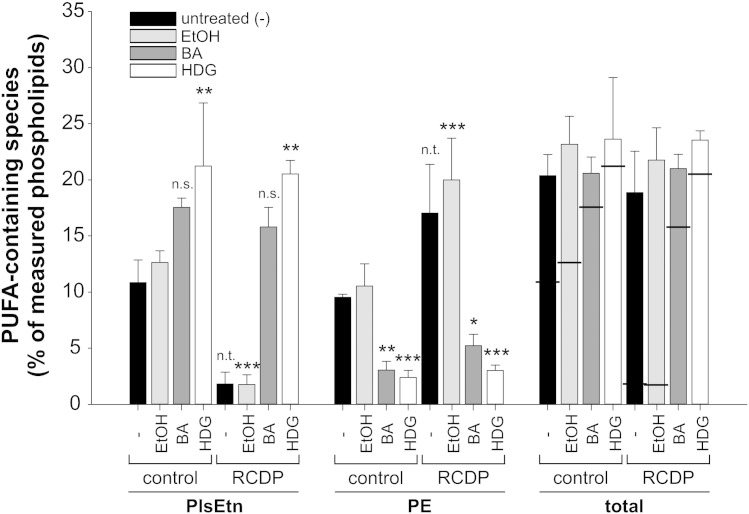
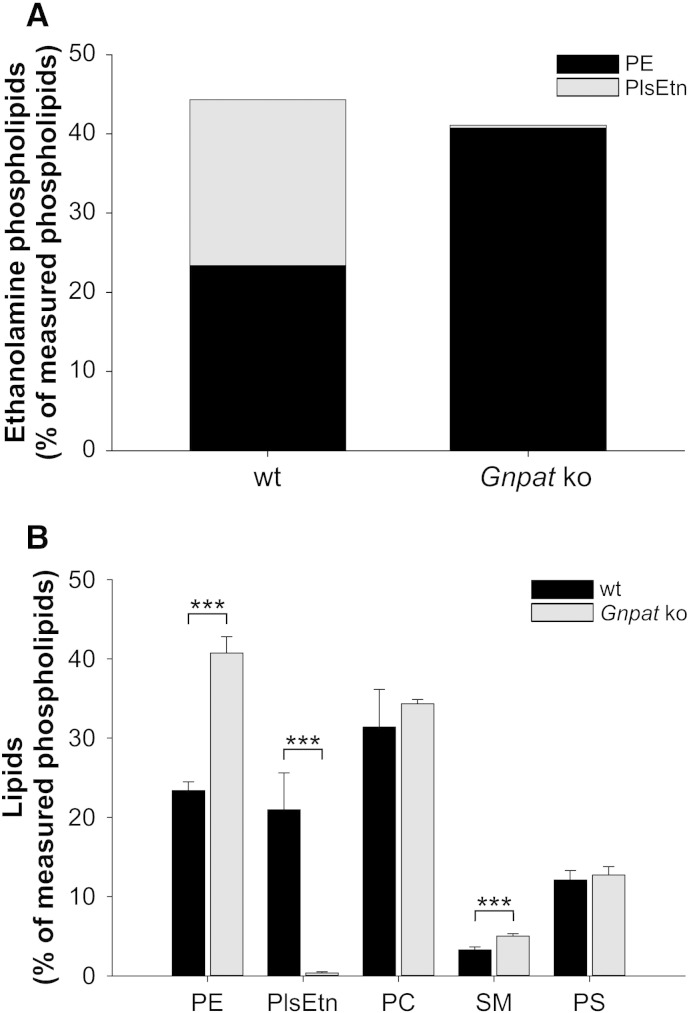
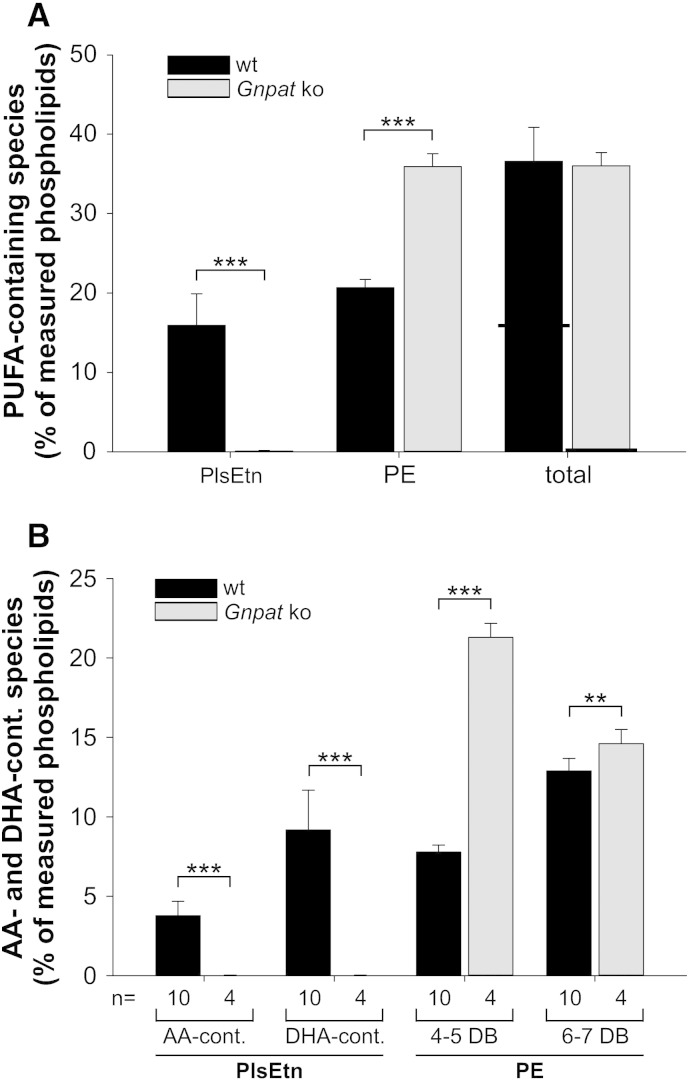
Ethanolamine plasmalogens constitute a group of ether glycerophospholipids that, due to their unique biophysical and biochemical properties, are essential components of mammalian cellular membranes. Their importance is emphasized by the consequences of defects in plasmalogen biosynthesis, which in humans cause the fatal disease rhizomelic chondrodysplasia punctata (RCDP). In the present lipidomic study, we used fibroblasts derived from RCDP patients, as well as brain tissue from plasmalogen-deficient mice, to examine the compensatory mechanisms of lipid homeostasis in response to plasmalogen deficiency. Our results show that phosphatidylethanolamine (PE), a diacyl glycerophospholipid, which like ethanolamine plasmalogens carries the head group ethanolamine, is the main player in the adaptation to plasmalogen insufficiency. PE levels were tightly adjusted to the amount of ethanolamine plasmalogens so that their combined levels were kept constant. Similarly, the total amount of polyunsaturated fatty acids (PUFAs) in ethanolamine phospholipids was maintained upon plasmalogen deficiency. However, we found an increased incorporation of arachidonic acid at the expense of docosahexaenoic acid in the PE fraction of plasmalogen-deficient tissues. These data show that under conditions of reduced plasmalogen levels, the amount of total ethanolamine phospholipids is precisely maintained by a rise in PE. At the same time, a shift in the ratio between ω-6 and ω-3 PUFAs occurs, which might have unfavorable, long-term biological consequences. Therefore, our findings are not only of interest for RCDP but may have more widespread implications also for other disease conditions, as for example Alzheimer's disease, that have been associated with a decline in plasmalogens.
Keywords: Alzheimer's disease; Arachidonic acid; Compensation; Docosahexaenoic acid; Peroxisome; Plasmalogen.
Copyright © 2014. Published by Elsevier B.V.
Figures
References
-
- Coskun U., Simons K. Cell membranes: the lipid perspective. Structure. 2011;19:1543–1548. - PubMed
-
- Hazel J.R., Williams E.E. The role of alterations in membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 1990;29:167–227. - PubMed
-
- Guido M.E., Garbarino Pico E., Caputto B.L. Circadian regulation of phospholipid metabolism in retinal photoreceptors and ganglion cells. J. Neurochem. 2001;76:835–845. - PubMed
-
- Pike L.J. Rafts defined: a report on the Keystone symposium on lipid rafts and cell function. J. Lipid Res. 2006;47:1597–1598. - PubMed
-
- Nagan N., Zoeller R.A. Plasmalogens: biosynthesis and functions. Prog. Lipid Res. 2001;40:199–229. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
