

Osteoblast-specific deletion of Pkd2 leads to low-turnover osteopenia and reduced bone marrow adiposity
- PMID: 25464512
- PMCID: PMC4252138
- DOI: 10.1371/journal.pone.0114198
Osteoblast-specific deletion of Pkd2 leads to low-turnover osteopenia and reduced bone marrow adiposity
Abstract
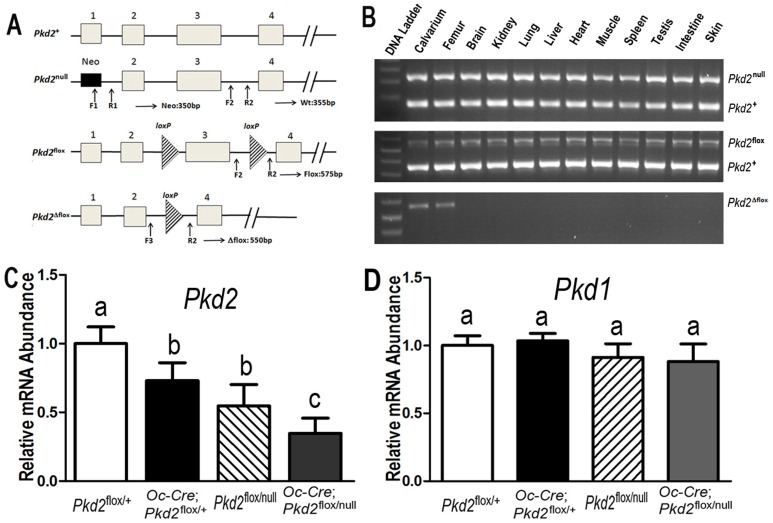
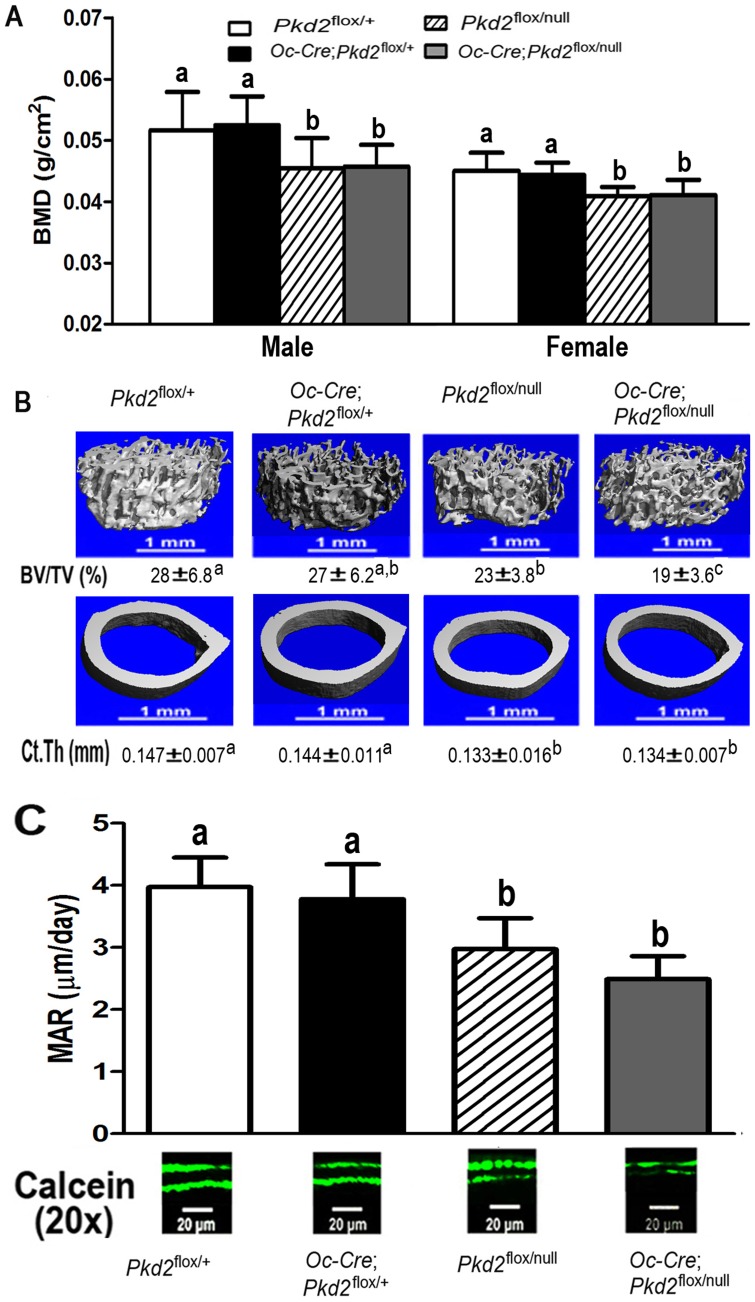
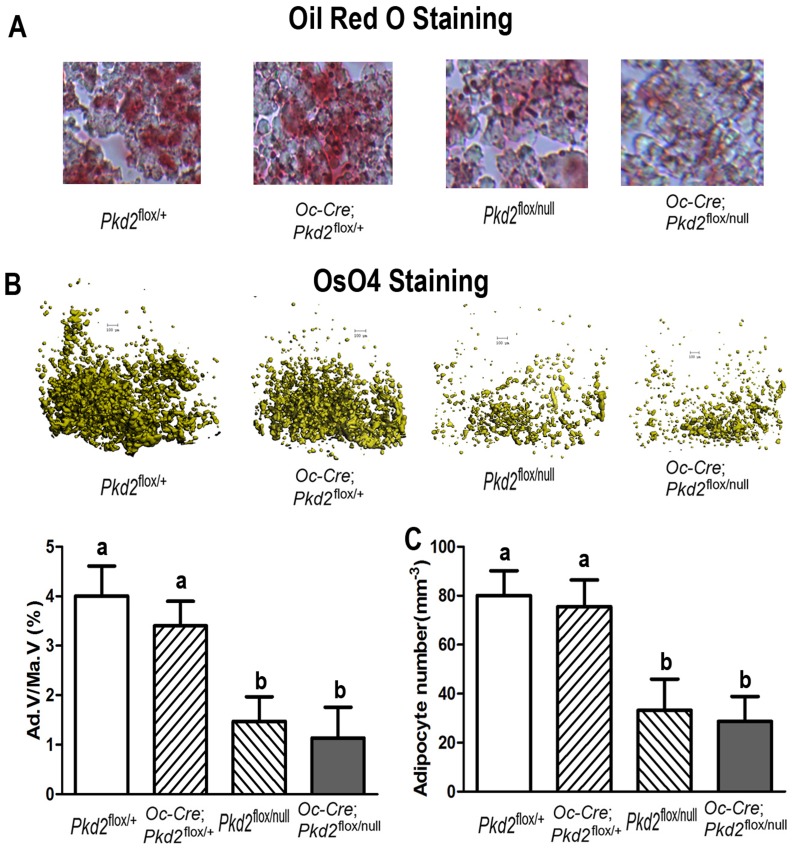
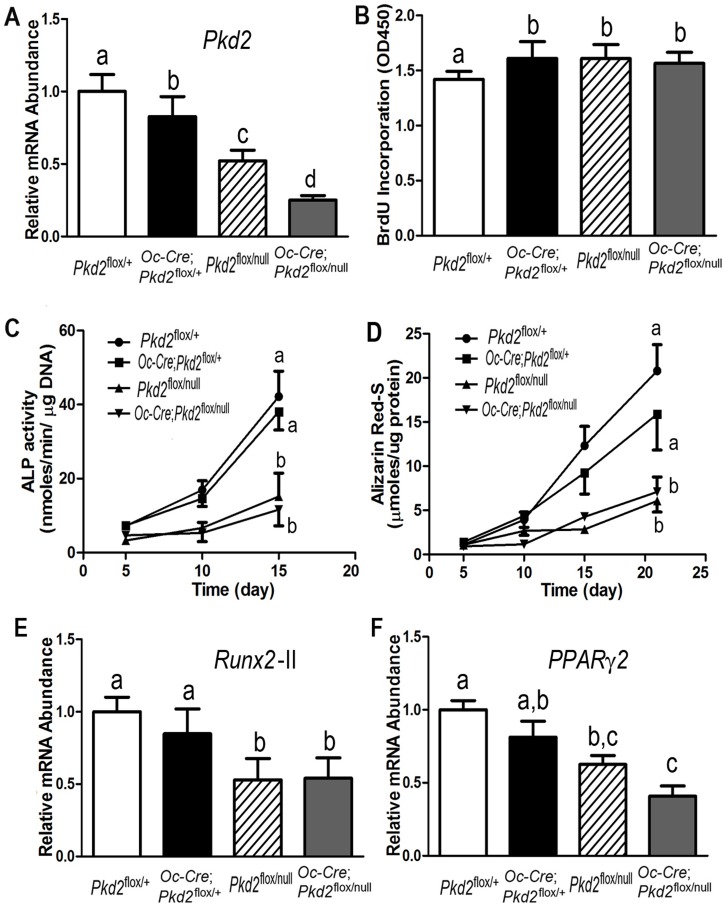
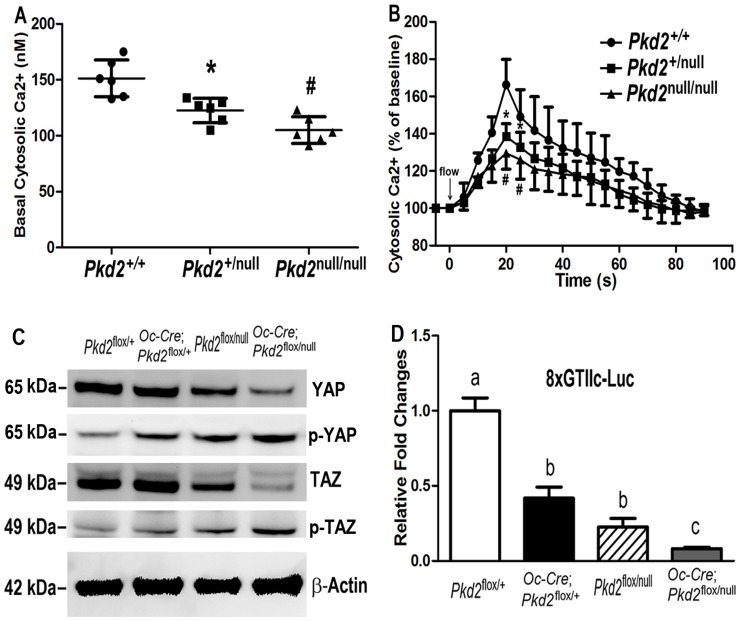
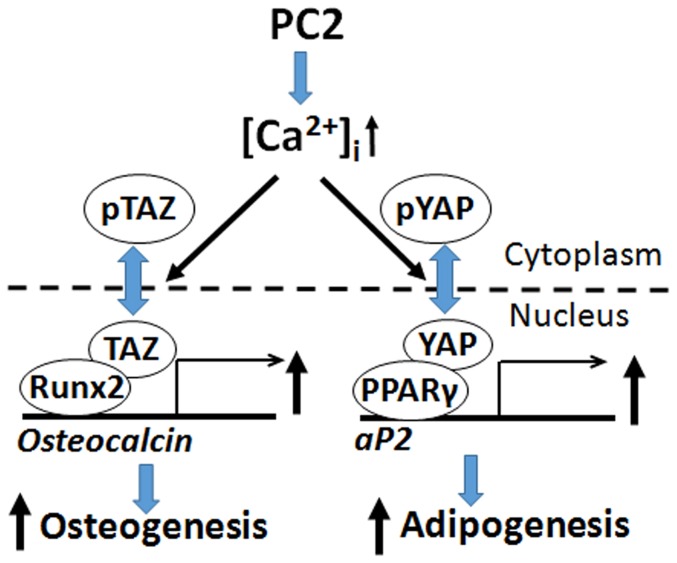
Polycystin-1 (Pkd1) interacts with polycystin-2 (Pkd2) to form an interdependent signaling complex. Selective deletion of Pkd1 in the osteoblast lineage reciprocally regulates osteoblastogenesis and adipogenesis. The role of Pkd2 in skeletal development has not been defined. To this end, we conditionally inactivated Pkd2 in mature osteoblasts by crossing Osteocalcin (Oc)-Cre;Pkd2+/null mice with floxed Pkd2 (Pkd2flox/flox) mice. Oc-Cre;Pkd2flox/null (Pkd2Oc-cKO) mice exhibited decreased bone mineral density, trabecular bone volume, cortical thickness, mineral apposition rate and impaired biomechanical properties of bone. Pkd2 deficiency resulted in diminished Runt-related transcription factor 2 (Runx2) expressions in bone and impaired osteoblastic differentiation ex vivo. Expression of osteoblast-related genes, including, Osteocalcin, Osteopontin, Bone sialoprotein (Bsp), Phosphate-regulating gene with homologies to endopeptidases on the X chromosome (Phex), Dentin matrix protein 1 (Dmp1), Sclerostin (Sost), and Fibroblast growth factor 23 (FGF23) were reduced proportionate to the reduction of Pkd2 gene dose in bone of Oc-Cre;Pkd2flox/+ and Oc-Cre;Pkd2flox/null mice. Loss of Pkd2 also resulted in diminished peroxisome proliferator-activated receptor γ (PPARγ) expression and reduced bone marrow fat in vivo and reduced adipogenesis in osteoblast culture ex vivo. Transcriptional co-activator with PDZ-binding motif (TAZ) and Yes-associated protein (YAP), reciprocally acting as co-activators and co-repressors of Runx2 and PPARγ, were decreased in bone of Oc-Cre;Pkd2flox/null mice. Thus, Pkd1 and Pkd2 have coordinate effects on osteoblast differentiation and opposite effects on adipogenesis, suggesting that Pkd1 and Pkd2 signaling pathways can have independent effects on mesenchymal lineage commitment in bone.
Conflict of interest statement
Figures
References
-
- Qian F, Germino FJ, Cai Y, Zhang X, Somlo S, et al. (1997) PKD1 interacts with PKD2 through a probable coiled-coil domain. Nat Genet 16:179–183. - PubMed
-
- Hanaoka K, Qian F, Boletta A, Bhunia AK, Piontek K, et al. (2000) Co-assembly of polycystin-1 and -2 produces unique cation-permeable currents. Nature 408:990–994. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
