

Arabinogalactan proteins are involved in root hair development in barley
- PMID: 25465033
- PMCID: PMC4339589
- DOI: 10.1093/jxb/eru475
Arabinogalactan proteins are involved in root hair development in barley
Abstract
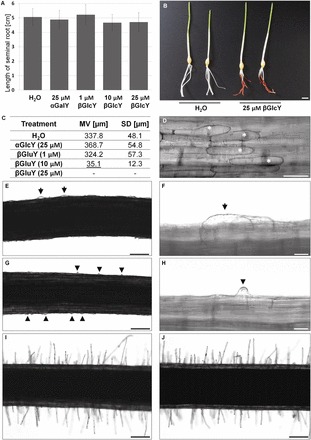
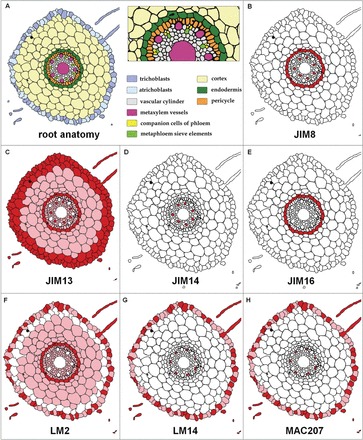
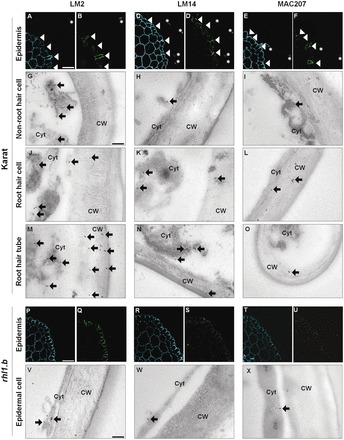
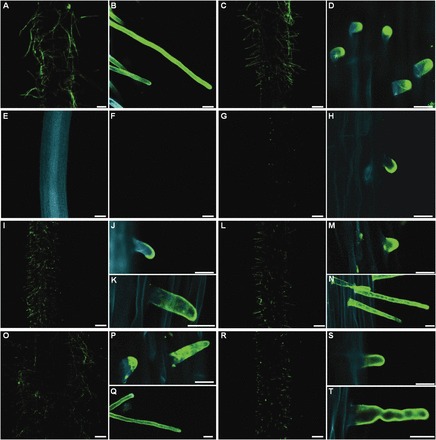
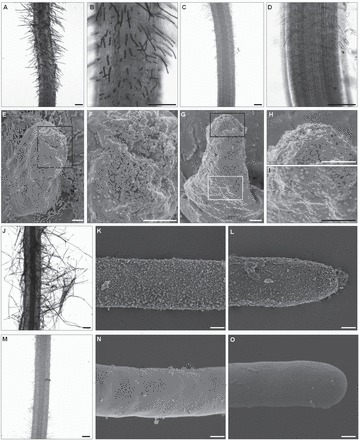
The arabinogalactan proteins (AGPs) are involved in a range of plant processes, including cell differentiation and expansion. Here, barley root hair mutants and their wild-type parent cultivars were used, as a model system, to reveal the role of AGPs in root hair development. The treatment of roots with different concentrations of βGlcY (a reagent which binds to all classes of AGPs) inhibited or totally suppressed the development of root hairs in all of the cultivars. Three groups of AGP (recognized by the monoclonal antibodies LM2, LM14, and MAC207) were diversely localized in trichoblasts and atrichoblasts of root hair-producing plants. The relevant epitopes were present in wild-type trichoblast cell walls and cytoplasm, whereas in wild-type atrichoblasts and in all epidermal cells of a root hairless mutant, they were only present in the cytoplasm. In all of cultivars the higher expression of LM2, LM14, and MAC207 was observed in trichoblasts at an early stage of development. Additionally, the LM2 epitope was detected on the surface of primordia and root hair tubes in plants able to generate root hairs. The major conclusion was that the AGPs recognized by LM2, LM14, and MAC207 are involved in the differentiation of barley root epidermal cells, thereby implying a requirement for these AGPs for root hair development in barley.
Keywords: Arabinogalactan proteins (AGPs); Yariv.; barley (Hordeum vulgare); cell differentiation; monoclonal antibodies; root hairs.
© The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Andeme-Onzighi C, Sivaguru M, Judy-March J, Baskin TI, Driouich A. 2002. The reb1-1 mutation of Arabidopsis alters the morphology of trichoblasts, the expression of arabinogalactan proteins and the organisation of cortical microtubules. Planta 215, 949–958. - PubMed
-
- Bradley DJ, Wood EA, Larkins AP, Galfre G, Butcher GW, Brewin NJ. 1988. Isolation of monoclonal antibodies reacting with peribacteroid membranes and other components of pea root nodules containing Rhizobium leguminosarum . Planta 173, 149–160. - PubMed
-
- Casero PJ, Casimiro I, Knox JP. 1998. Occurrence of cell surface arabinogalactan-protein and extensin epitopes in relation to pericycle and vascular tissue development in the root apex of four species. Planta 204, 252–259.
-
- Chen T, Teng N, Wu X, Wang Y, Tang W, Samaj J, Baluska F, Lin J. 2007. Disruption of actin filaments by latrunculin B affects cell wall construction in Picea meyeri pollen tube by disturbing vesicle trafficking. Plant and Cell Physiology 1, 19–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
