

Perivascular Gli1+ progenitors are key contributors to injury-induced organ fibrosis
- PMID: 25465115
- PMCID: PMC4289444
- DOI: 10.1016/j.stem.2014.11.004
Perivascular Gli1+ progenitors are key contributors to injury-induced organ fibrosis
Abstract
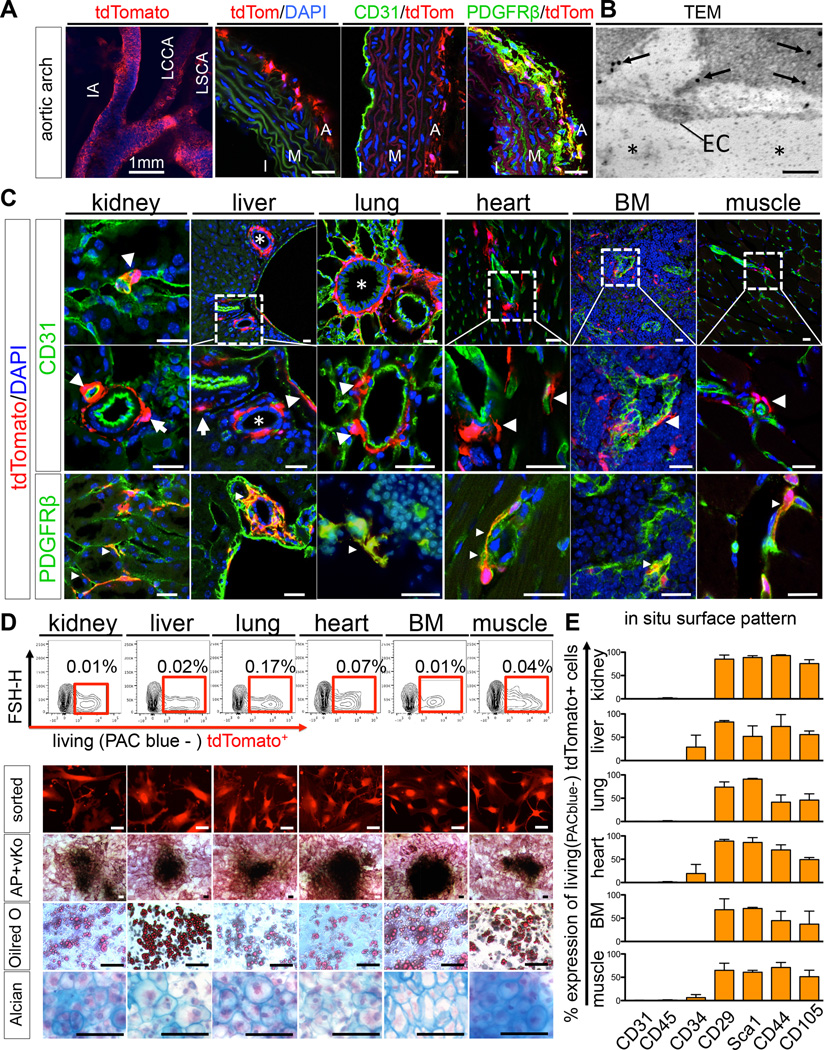
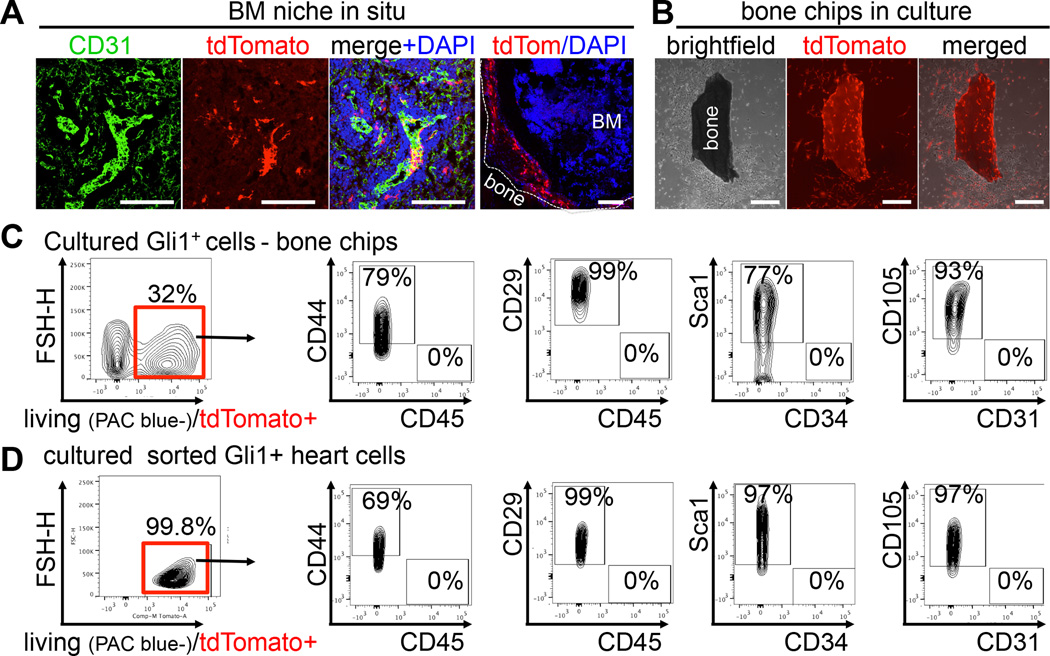
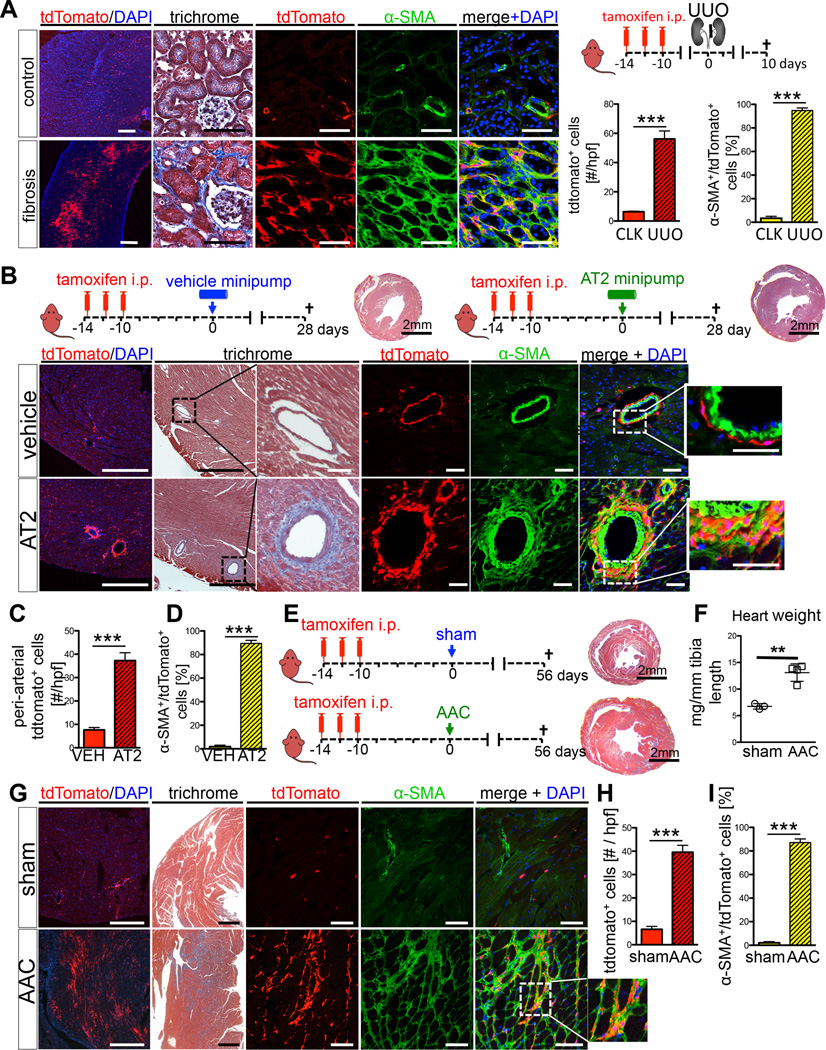
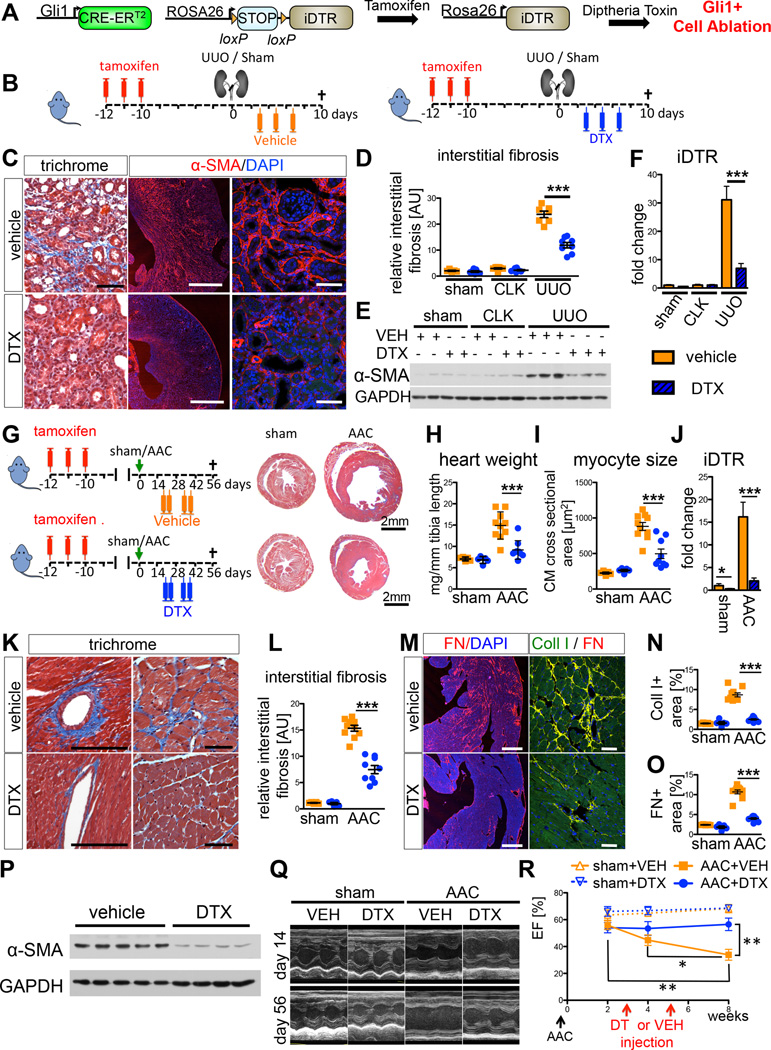
Mesenchymal stem cells (MSCs) reside in the perivascular niche of many organs, including kidney, lung, liver, and heart, although their roles in these tissues are poorly understood. Here, we demonstrate that Gli1 marks perivascular MSC-like cells that substantially contribute to organ fibrosis. In vitro, Gli1(+) cells express typical MSC markers, exhibit trilineage differentiation capacity, and possess colony-forming activity, despite constituting a small fraction of the platelet-derived growth factor-β (PDGFRβ)(+) cell population. Genetic lineage tracing analysis demonstrates that tissue-resident, but not circulating, Gli1(+) cells proliferate after kidney, lung, liver, or heart injury to generate myofibroblasts. Genetic ablation of these cells substantially ameliorates kidney and heart fibrosis and preserves ejection fraction in a model of induced heart failure. These findings implicate perivascular Gli1(+) MSC-like cells as a major cellular origin of organ fibrosis and demonstrate that these cells may be a relevant therapeutic target to prevent solid organ dysfunction after injury.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures



Comment in
-
Targeting the cellular origin of organ fibrosis.Cell Stem Cell. 2015 Jan 8;16(1):3-4. doi: 10.1016/j.stem.2014.12.008. Cell Stem Cell. 2015. PMID: 25575074
References
-
- Caplan AI. Mesenchymal stem cells. Journal of orthopaedic research : official publication of the Orthopaedic Research Society. 1991;9:641–650. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
