

Regulation of kinetochore-microtubule attachments through homeostatic control during mitosis
- PMID: 25466864
- PMCID: PMC4568440
- DOI: 10.1038/nrm3916
Regulation of kinetochore-microtubule attachments through homeostatic control during mitosis
Abstract
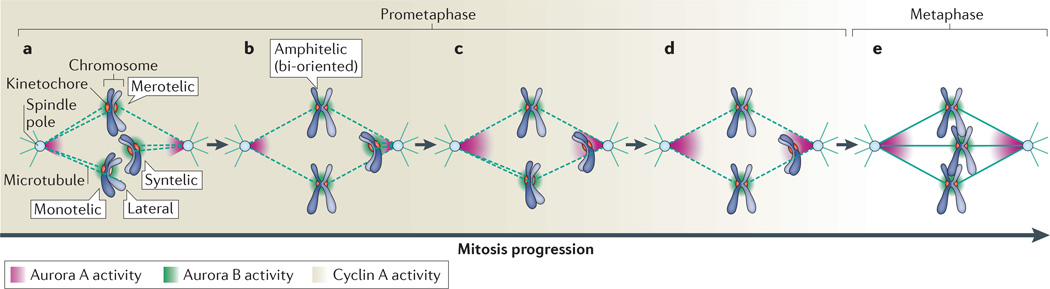
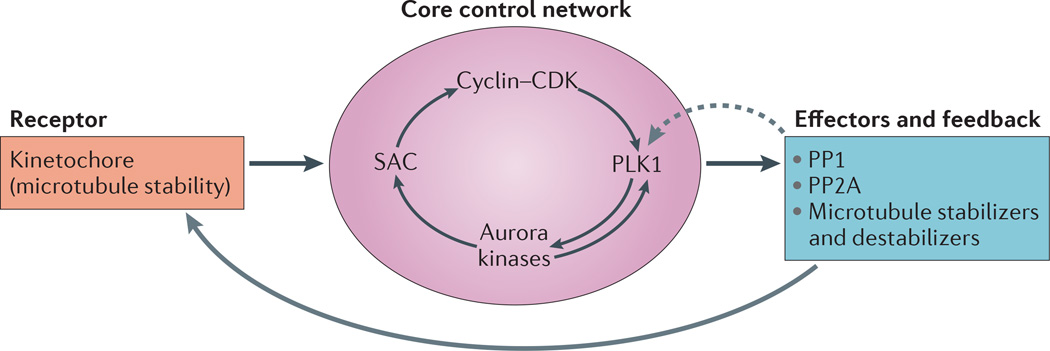
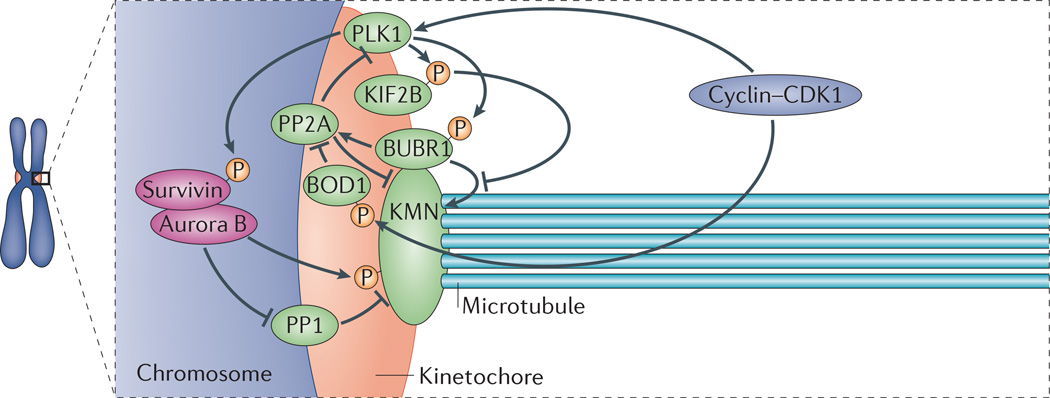
Faithful chromosome segregation during mitosis is essential for genome integrity and is mediated by the bi-oriented attachment of replicated chromosomes to spindle microtubules through kinetochores. Errors in kinetochore-microtubule (k-MT) attachment that could cause chromosome mis-segregation are frequent and are corrected by the dynamic turnover of k-MT attachments. Thus, regulating the rate of spindle microtubule attachment and detachment to kinetochores is crucial for mitotic fidelity and is frequently disrupted in cancer cells displaying chromosomal instability. A model based on homeostatic principles involving receptors, a core control network, effectors and feedback control may explain the precise regulation of k-MT attachment stability during mitotic progression to ensure error-free mitosis.
Figures
References
-
- Cimini D, Degrassi F. Aneuploidy: a matter of bad connections. Trends Cell Biol. 2001;15:442–451. - PubMed
-
- Cimini D. Merotelic kinetochore orientation, aneuploidy, and cancer. Biochim. Biophys. Acta. 2008;1786:32–40. - PubMed
-
- Duijf PH, Benezra R. The cancer biology of whole-chromosome instability. Oncogene. 2013;32:4727–4736. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
