

Microtubule nucleation remote from centrosomes may explain how asters span large cells
- PMID: 25468969
- PMCID: PMC4273342
- DOI: 10.1073/pnas.1418796111
Microtubule nucleation remote from centrosomes may explain how asters span large cells
Abstract
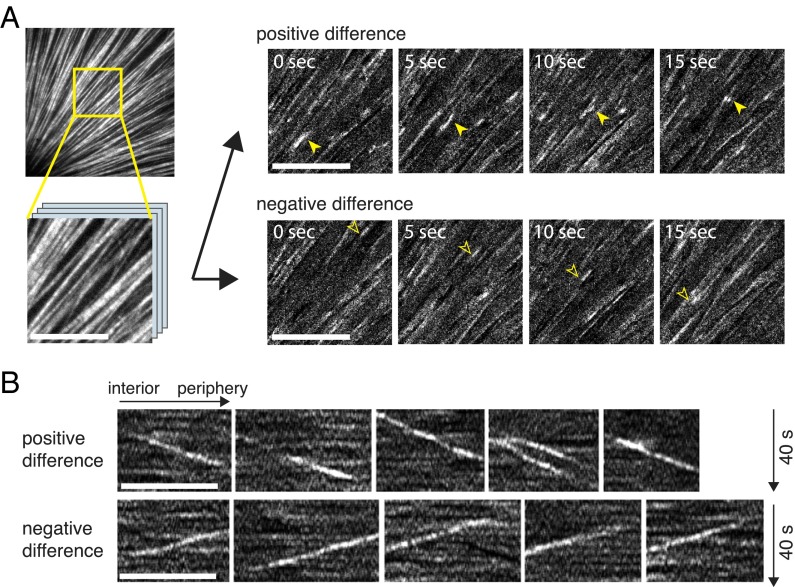
A major challenge in cell biology is to understand how nanometer-sized molecules can organize micrometer-sized cells in space and time. One solution in many animal cells is a radial array of microtubules called an aster, which is nucleated by a central organizing center and spans the entire cytoplasm. Frog (here Xenopus laevis) embryos are more than 1 mm in diameter and divide with a defined geometry every 30 min. Like smaller cells, they are organized by asters, which grow, interact, and move to precisely position the cleavage planes. It has been unclear whether asters grow to fill the enormous egg by the same mechanism used in smaller somatic cells, or whether special mechanisms are required. We addressed this question by imaging growing asters in a cell-free system derived from eggs, where asters grew to hundreds of microns in diameter. By tracking marks on the lattice, we found that microtubules could slide outward, but this was not essential for rapid aster growth. Polymer treadmilling did not occur. By measuring the number and positions of microtubule ends over time, we found that most microtubules were nucleated away from the centrosome and that interphase egg cytoplasm supported spontaneous nucleation after a time lag. We propose that aster growth is initiated by centrosomes but that asters grow by propagating a wave of microtubule nucleation stimulated by the presence of preexisting microtubules.
Keywords: Xenopus; aster; centrosome; embryo; microtubule nucleation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Wilson EB. The Cell in Development and Inheritance. Macmillan; New York: 1925.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
