

MapZ marks the division sites and positions FtsZ rings in Streptococcus pneumoniae
- PMID: 25470041
- PMCID: PMC4268495
- DOI: 10.1038/nature13966
MapZ marks the division sites and positions FtsZ rings in Streptococcus pneumoniae
Abstract
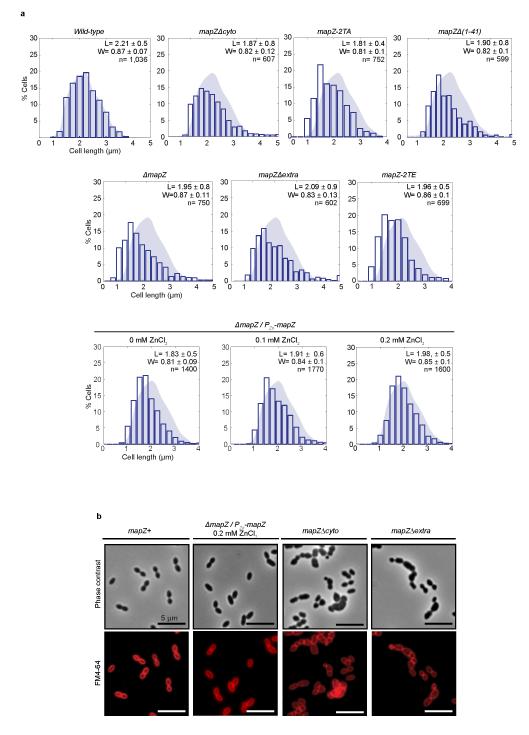
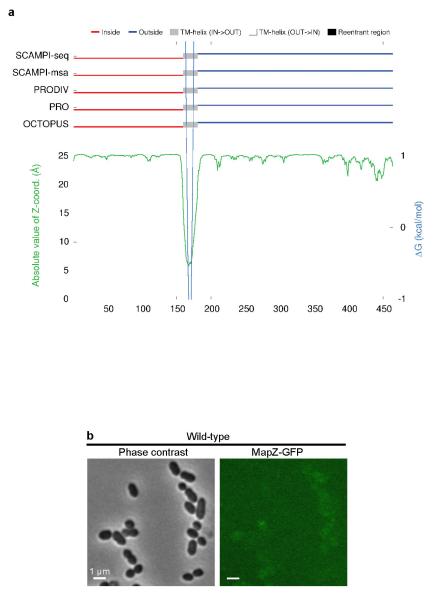
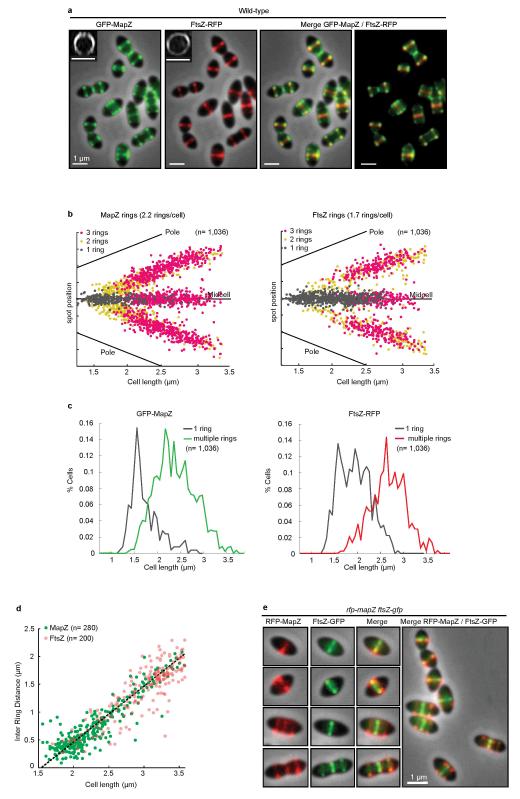
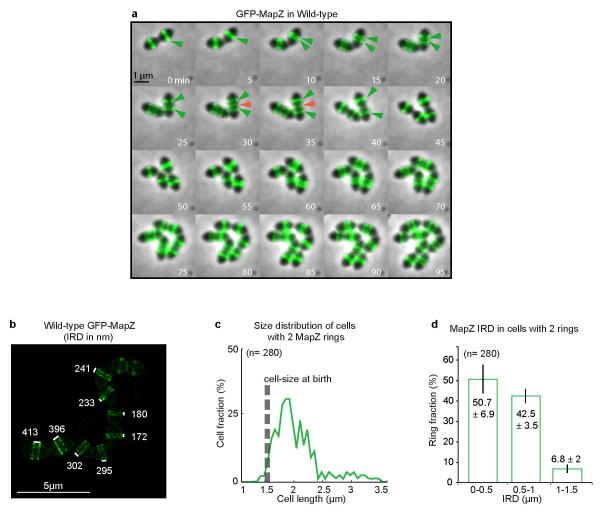
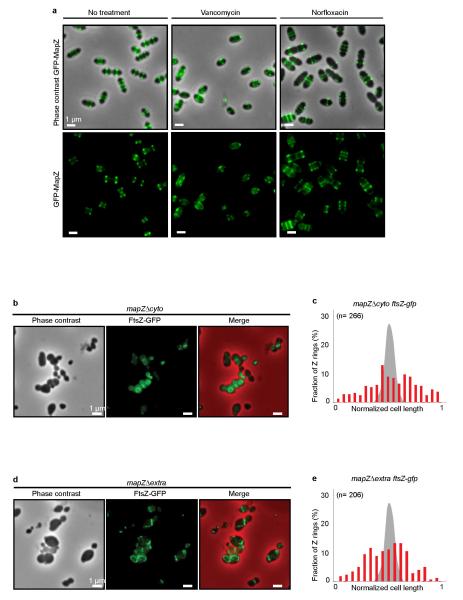
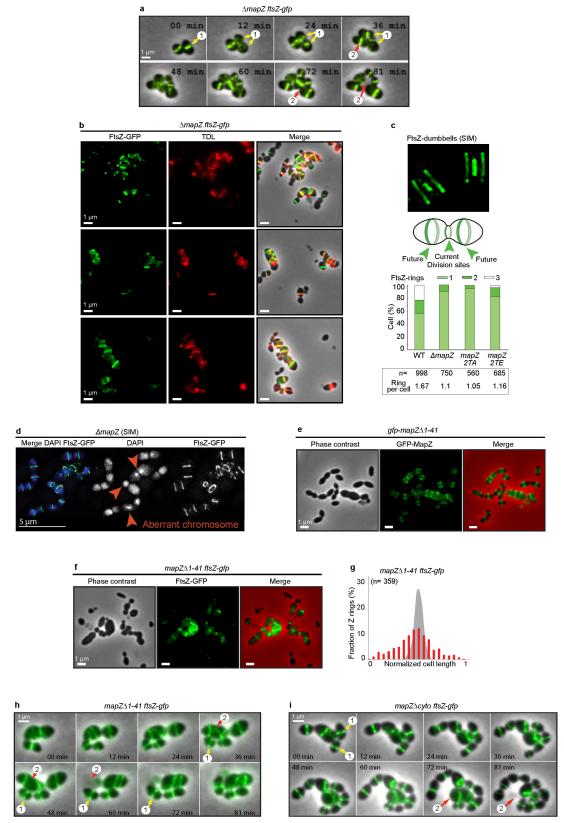
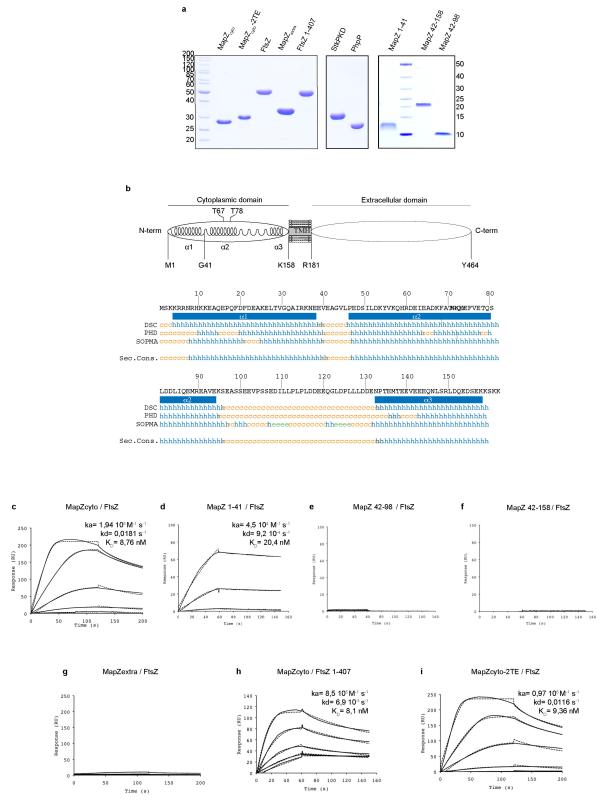
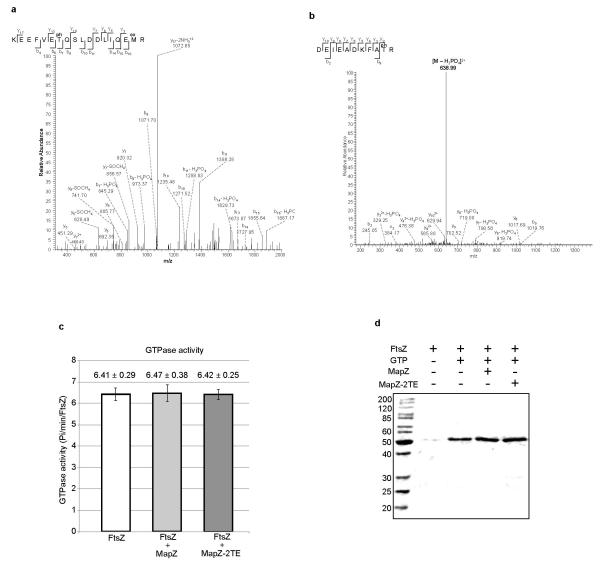
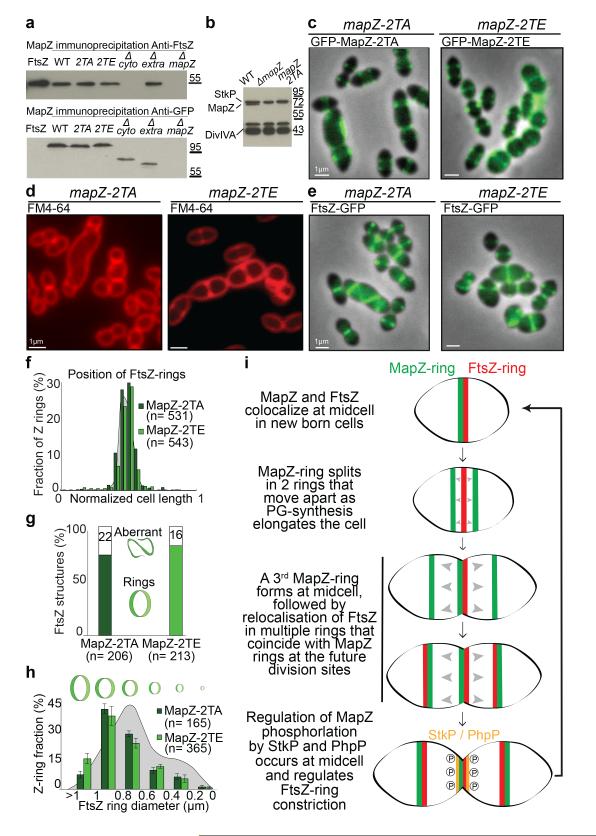
In every living organism, cell division requires accurate identification of the division site and placement of the division machinery. In bacteria, this process is traditionally considered to begin with the polymerization of the highly conserved tubulin-like protein FtsZ into a ring that locates precisely at mid-cell. Over the past decades, several systems have been reported to regulate the spatiotemporal assembly and placement of the FtsZ ring. However, the human pathogen Streptococcus pneumoniae, in common with many other organisms, is devoid of these canonical systems and the mechanisms of positioning the division machinery remain unknown. Here we characterize a novel factor that locates at the division site before FtsZ and guides septum positioning in pneumococcus. Mid-cell-anchored protein Z (MapZ) forms ring structures at the cell equator and moves apart as the cell elongates, therefore behaving as a permanent beacon of division sites. MapZ then positions the FtsZ ring through direct protein-protein interactions. MapZ-mediated control differs from previously described systems mostly on the basis of negative regulation of FtsZ assembly. Furthermore, MapZ is an endogenous target of the Ser/Thr kinase StkP, which was recently shown to have a central role in cytokinesis and morphogenesis of S. pneumoniae. We show that both phosphorylated and non-phosphorylated forms of MapZ are required for proper Z-ring formation and dynamics. Altogether, this work uncovers a new mechanism for bacterial cell division that is regulated by phosphorylation and illustrates that nature has evolved a diversity of cell division mechanisms adapted to the different bacterial clades.
Figures
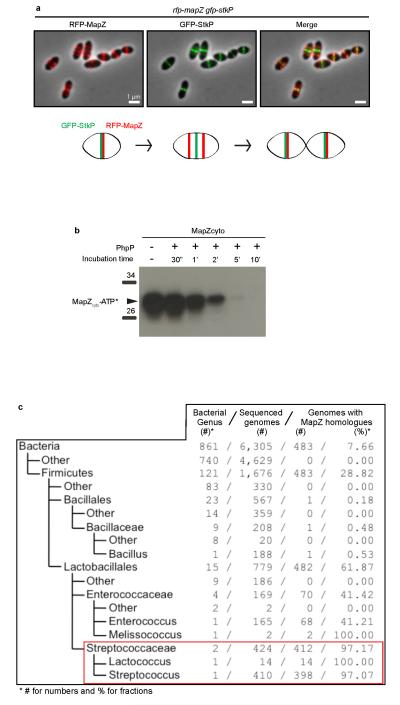
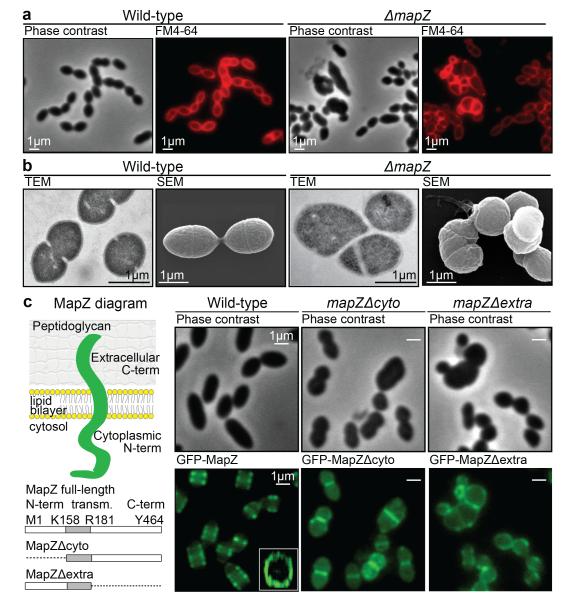
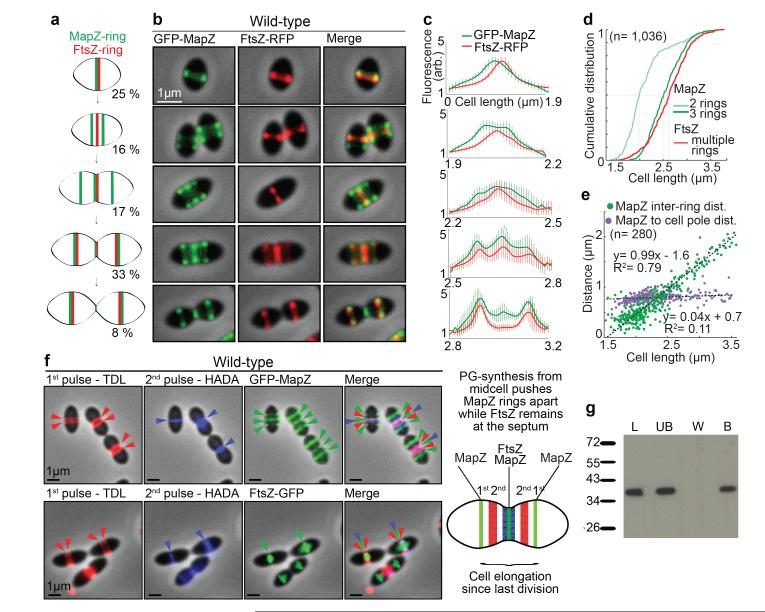
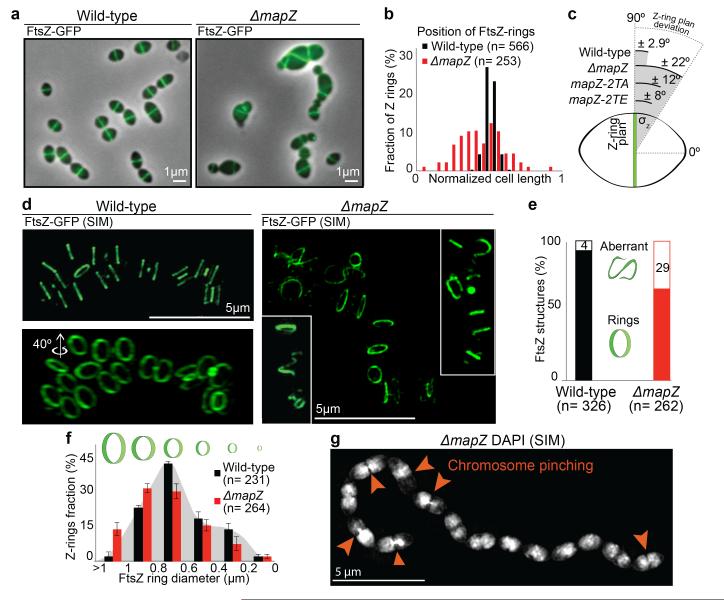
Comment in
-
Microbiology: A beacon for bacterial tubulin.Nature. 2014 Dec 11;516(7530):175-6. doi: 10.1038/nature14071. Epub 2014 Nov 26. Nature. 2014. PMID: 25470050 No abstract available.
-
Following the equator: division site selection in Streptococcus pneumoniae.Trends Microbiol. 2015 Mar;23(3):121-2. doi: 10.1016/j.tim.2015.02.001. Epub 2015 Feb 12. Trends Microbiol. 2015. PMID: 25684260
-
Mapping mid-cell: MapZ shows the way.Cell Cycle. 2015;14(7):937-8. doi: 10.1080/15384101.2015.1010978. Cell Cycle. 2015. PMID: 25803432 Free PMC article. No abstract available.
References
-
- Adams DW, Errington J. Bacterial cell division: assembly, maintenance and disassembly of the Z ring. Nat Rev Microbiol. 2009;7:642–653. doi:10.1038/nrmicro2198. - PubMed
-
- Bramkamp M, van Baarle S. Division site selection in rod-shaped bacteria. Curr Opin Microbiol. 2009;12:683–688. doi:10.1016/j.mib.2009.10.002. - PubMed
-
- Wu LJ, Errington J. Nucleoid occlusion and bacterial cell division. Nat Rev Microbiol. 2012;10:8–12. doi:10.1038/nrmicro2671. - PubMed
-
- Pinho MG, Kjos M, Veening JW. How to get (a)round: mechanisms controlling growth and division of coccoid bacteria. Nat Rev Microbiol. 2013;11:601–614. doi:10.1038/nrmicro3088. - PubMed
-
- Monahan LG, Harry EJ. Identifying how bacterial cells find their middle: a new perspective. Mol Microbiol. 2013;87:231–234. doi:10.1111/mmi.12114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
