

A parabrachial-hypothalamic cholecystokinin neurocircuit controls counterregulatory responses to hypoglycemia
- PMID: 25470549
- PMCID: PMC4261079
- DOI: 10.1016/j.cmet.2014.11.006
A parabrachial-hypothalamic cholecystokinin neurocircuit controls counterregulatory responses to hypoglycemia
Abstract
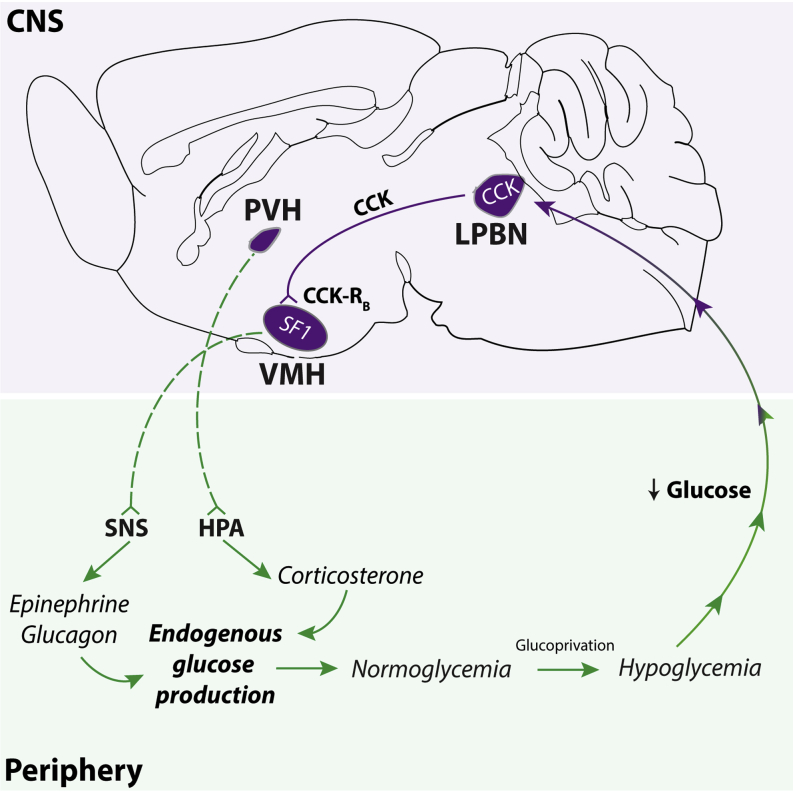
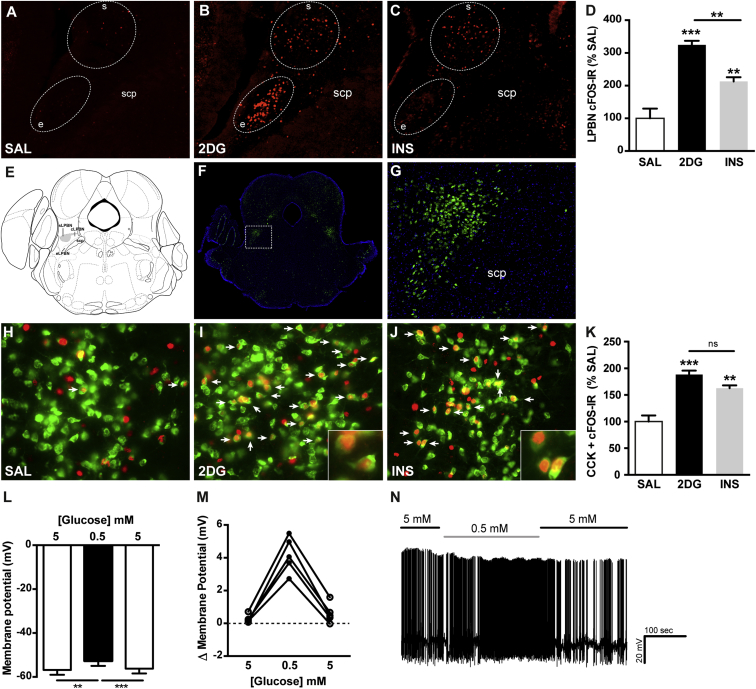
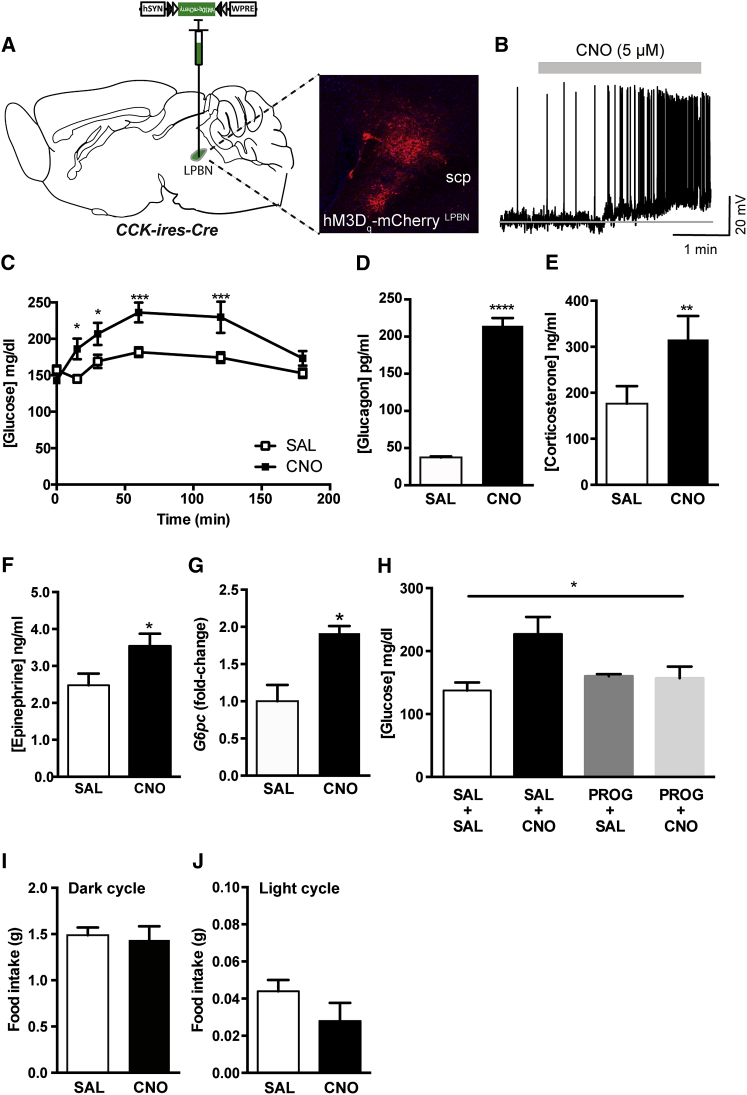
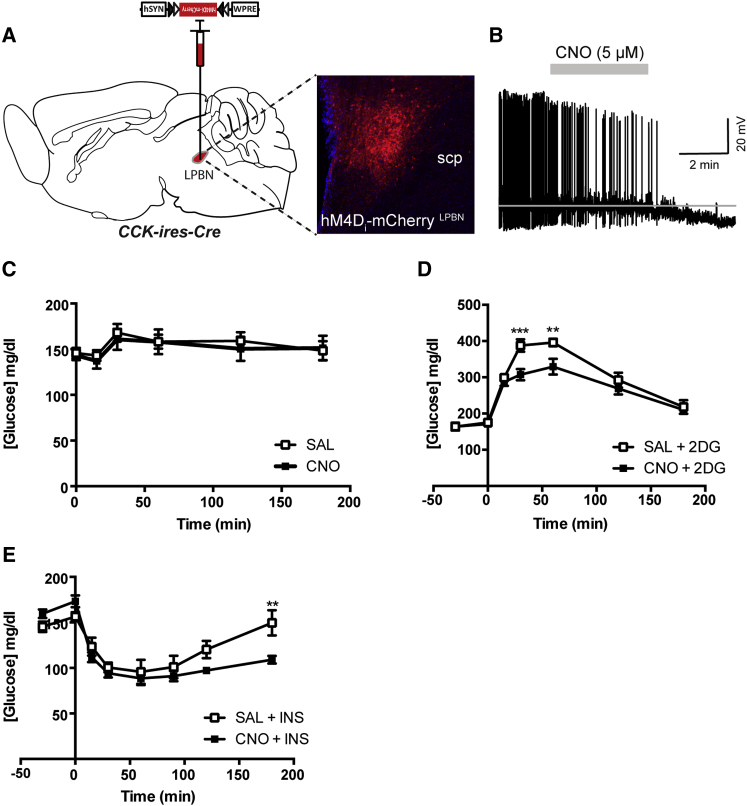
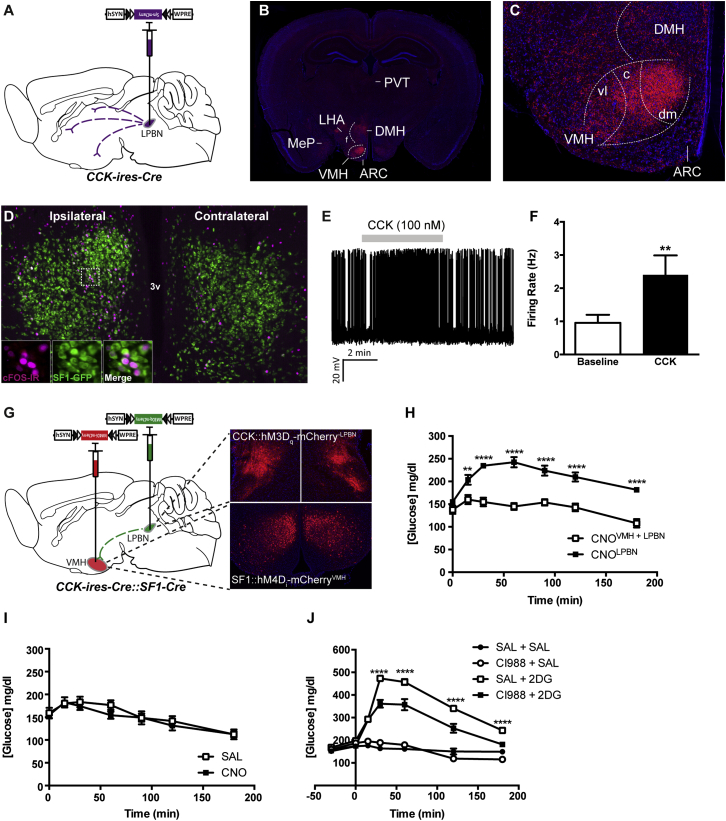
Hypoglycemia engenders an autonomically mediated counterregulatory (CR)-response that stimulates endogenous glucose production to maintain concentrations within an appropriate physiological range. Although the involvement of the brain in preserving normoglycemia has been established, the neurocircuitry underlying centrally mediated CR-responses remains unclear. Here we demonstrate that lateral parabrachial nucleus cholecystokinin (CCK(LPBN)) neurons are a population of glucose-sensing cells (glucose inhibited) with counterregulatory capacity. Furthermore, we reveal that steroidogenic-factor 1 (SF1)-expressing neurons of the ventromedial nucleus of the hypothalamus (SF1(VMH)) are the specific target of CCK(LPBN) glucoregulatory neurons. This discrete CCK(LPBN)→SF1(VMH) neurocircuit is both necessary and sufficient for the induction of CR-responses. Together, these data identify CCK(LPBN) neurons, and specifically CCK neuropeptide, as glucoregulatory and provide significant insight into the homeostatic mechanisms controlling CR-responses to hypoglycemia.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Expanding the brain glucosensing territory.Cell Metab. 2014 Dec 2;20(6):933-5. doi: 10.1016/j.cmet.2014.11.013. Cell Metab. 2014. PMID: 25470543
References
-
- Borg W.P., Sherwin R.S., During M.J., Borg M.A., Shulman G.I. Local ventromedial hypothalamus glucopenia triggers counterregulatory hormone release. Diabetes. 1995;44:180–184. - PubMed
-
- Briski K.P. Induction of Fos immunoreactivity by acute glucose deprivation in the rat caudal brainstem: relation to NADPH diaphorase localization. Histochem. Cell Biol. 1999;111:229–233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK089044/DK/NIDDK NIH HHS/United States
- DK059637/DK/NIDDK NIH HHS/United States
- DK020593/DK/NIDDK NIH HHS/United States
- R01 DK098853/DK/NIDDK NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- R01 DK075632/DK/NIDDK NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- U24 DK059637/DK/NIDDK NIH HHS/United States
- R01 DK096010/DK/NIDDK NIH HHS/United States
- R37 DK053477/DK/NIDDK NIH HHS/United States
- R01 DK071051/DK/NIDDK NIH HHS/United States
- 098012/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
