

Cytomegalovirus-mediated activation of pyrimidine biosynthesis drives UDP-sugar synthesis to support viral protein glycosylation
- PMID: 25472841
- PMCID: PMC4273352
- DOI: 10.1073/pnas.1415864111
Cytomegalovirus-mediated activation of pyrimidine biosynthesis drives UDP-sugar synthesis to support viral protein glycosylation
Abstract
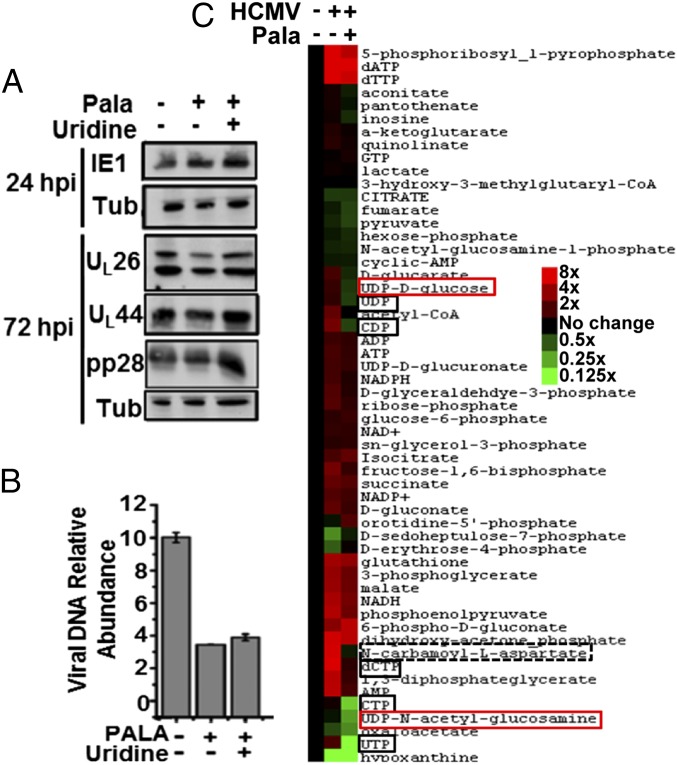
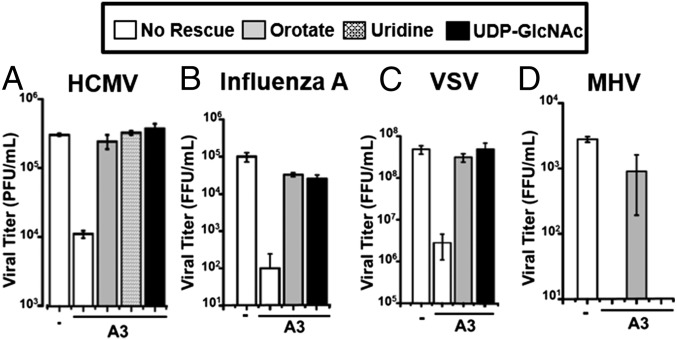
Human cytomegalovirus (HCMV) induces numerous changes to the host metabolic network that are critical for high-titer viral replication. We find that HCMV infection substantially induces de novo pyrimidine biosynthetic flux. This activation is important for HCMV replication because inhibition of pyrimidine biosynthetic enzymes substantially decreases the production of infectious virus, which can be rescued through medium supplementation with pyrimidine biosynthetic intermediates. Metabolomic analysis revealed that pyrimidine biosynthetic inhibition considerably reduces the levels of various UDP-sugar metabolites in HCMV-infected, but not mock-infected, cells. Further, UDP-sugar biosynthesis, which provides the sugar substrates required for glycosylation reactions, was found to be induced during HCMV infection. Pyrimidine biosynthetic inhibition also attenuated the glycosylation of the envelope glycoprotein B (gB). Both glycosylation of gB and viral growth were restored by medium supplementation with either UDP-sugar metabolites or pyrimidine precursors. These results indicate that HCMV drives de novo-synthesized pyrimidines to UDP-sugar biosynthesis to support virion protein glycosylation. The importance of this link between pyrimidine biosynthesis and UDP-sugars appears to be partially shared among diverse virus families, because UDP-sugar metabolites rescued the growth attenuation associated with pyrimidine biosynthetic inhibition during influenza A and vesicular stomatitis virus infection, but not murine hepatitis virus infection. In total, our results indicate that viruses can specifically modulate pyrimidine metabolic flux to provide the glycosyl subunits required for protein glycosylation and production of high titers of infectious progeny.
Keywords: UDP–sugar; cytomegalovirus; glycosylation; metabolism; pyrimidine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Bradley PL. Metabolism of pyruvate and alpha-ketoglutarate in virus-infected mouse brain. Nature. 1957;180(4599):1418–1419. - PubMed
-
- Kaplan AS, Ben-Porat T. The action of 5-fluorouracil on the nucleic acid metabolism of pseudorabies virus-infected and noninfected rabbit kidney cells. Virology. 1961;13:78–92. - PubMed
-
- Pearson HE, Winzler RJ. Oxidative and glycolytic metabolism of minced day-old mouse brain in relation to propagation of Theiler’s GD VII virus. J Biol Chem. 1949;181(2):577–582. - PubMed
-
- Goodrich JM, et al. Ganciclovir prophylaxis to prevent cytomegalovirus disease after allogeneic marrow transplant. Ann Intern Med. 1993;118(3):173–178. - PubMed
-
- Lee WA, Martin JC. Perspectives on the development of acyclic nucleotide analogs as antiviral drugs. Antiviral Res. 2006;71(2-3):254–259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R56 AI081773/AI/NIAID NIH HHS/United States
- R01AI081773/AI/NIAID NIH HHS/United States
- T32 AI049815/AI/NIAID NIH HHS/United States
- GM068411/GM/NIGMS NIH HHS/United States
- R03AI099681-01A1/AI/NIAID NIH HHS/United States
- R01 AI081773/AI/NIAID NIH HHS/United States
- T32 GM068411/GM/NIGMS NIH HHS/United States
- HHSN272201000055C/PHS HHS/United States
- HHSN266200700008C/AI/NIAID NIH HHS/United States
- HHSN266200700008C/PHS HHS/United States
- R03 AI099681/AI/NIAID NIH HHS/United States
- AI049815-13/AI/NIAID NIH HHS/United States
- HHSN272201000055C/AI/NIAID NIH HHS/United States
- R01 AI077719/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
