

Central nervous system regenerative failure: role of oligodendrocytes, astrocytes, and microglia
- PMID: 25475091
- PMCID: PMC4355267
- DOI: 10.1101/cshperspect.a020602
Central nervous system regenerative failure: role of oligodendrocytes, astrocytes, and microglia
Abstract
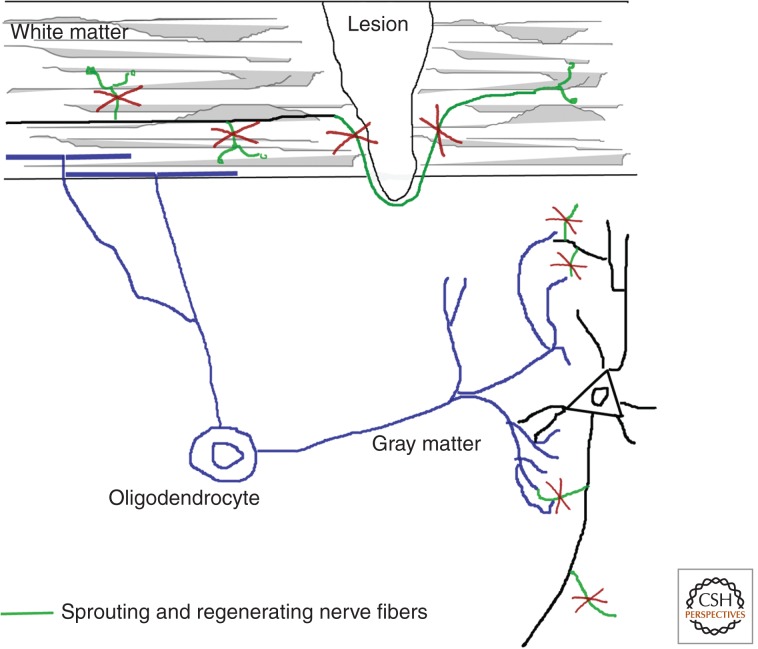
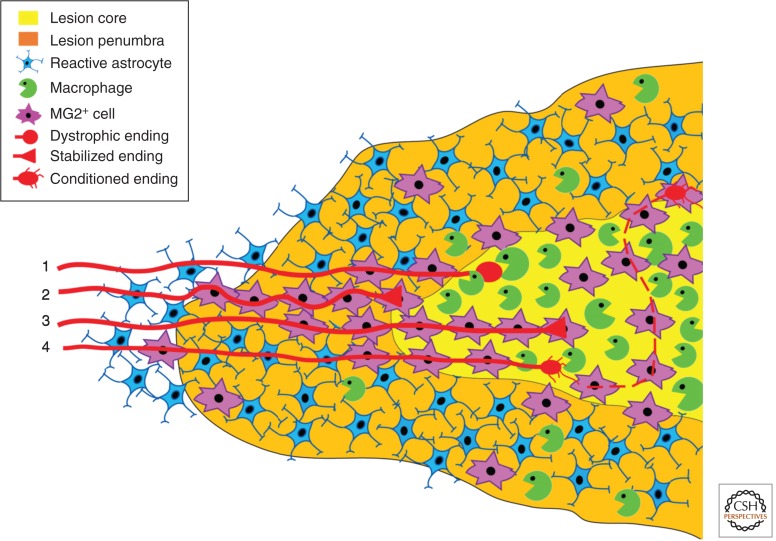
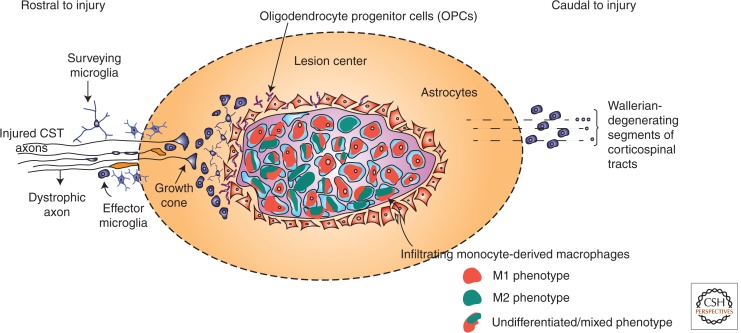
Animal studies are now showing the exciting potential to achieve significant functional recovery following central nervous system (CNS) injury by manipulating both the inefficient intracellular growth machinery in neurons, as well as the extracellular barriers, which further limit their regenerative potential. In this review, we have focused on the three major glial cell types: oligodendrocytes, astrocytes, and microglia/macrophages, in addition to some of their precursors, which form major extrinsic barriers to regrowth in the injured CNS. Although axotomized neurons in the CNS have, at best, a limited capacity to regenerate or sprout, there is accumulating evidence that even in the adult and, especially after boosting their growth motor, neurons possess the capacity for considerable circuit reorganization and even lengthy regeneration when these glial obstacles to neuronal regrowth are modified, eliminated, or overcome.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Abel R, Baron HC, Casha S, Harms J, Hurlbert J, Kucher K, Maier D, Thietje R, Weidner N, Curt A 2011. Therapeutic anti-Nogo-A antibodies in acute spinal cord injury: Safety and pharmacokinetic data from an ongoing first-in-human trial. ISCOS Meeting 2011, Washington, DC.
-
- Aguayo AJ, Rasminsky M, Bray GM, Carbonetto S, McKerracher L, Villegas-Prez M, Vidal-Sanz M, Carter DA 1991. Degenerative and regenerative responses of injured neurons in the central nervous system of adult mammals. Phil Trans R Soc B 331: 337–343. - PubMed
-
- Ahmed Z, Dent RG, Leadbeater WE, Smith C, Berry M, Logan A 2005. Matrix metalloproteases: Degradation of the inhibitory environment of the transected optic nerve and the scar by regenerating axons. Mol Cell Neurosci 28: 64–78. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical