

Effects of N-glycan precursor length diversity on quality control of protein folding and on protein glycosylation
- PMID: 25475176
- PMCID: PMC4452448
- DOI: 10.1016/j.semcdb.2014.11.008
Effects of N-glycan precursor length diversity on quality control of protein folding and on protein glycosylation
Abstract
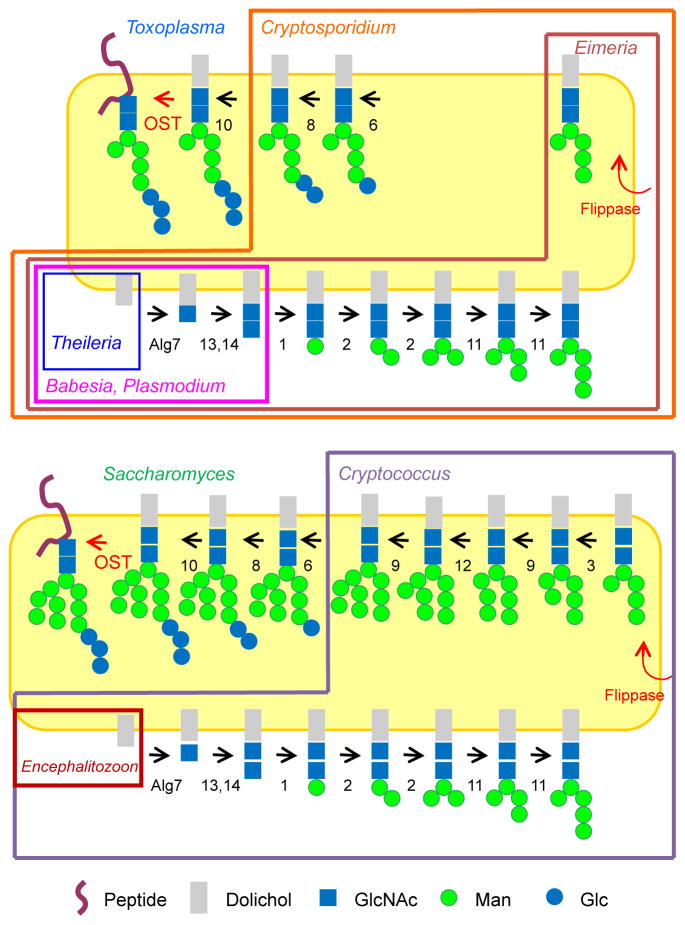
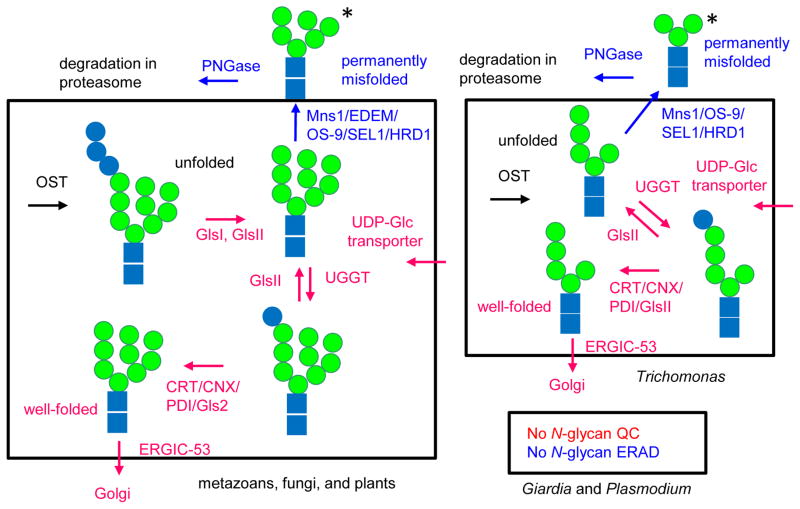
Asparagine-linked glycans (N-glycans) of medically important protists have much to tell us about the evolution of N-glycosylation and of N-glycan-dependent quality control (N-glycan QC) of protein folding in the endoplasmic reticulum. While host N-glycans are built upon a dolichol-pyrophosphate-linked precursor with 14 sugars (Glc3Man9GlcNAc2), protist N-glycan precursors vary from Glc3Man9GlcNAc2 (Acanthamoeba) to Man9GlcNAc2 (Trypanosoma) to Glc3Man5GlcNAc2 (Toxoplasma) to Man5GlcNAc2 (Entamoeba, Trichomonas, and Eimeria) to GlcNAc2 (Plasmodium and Giardia) to zero (Theileria). As related organisms have differing N-glycan lengths (e.g. Toxoplasma, Eimeria, Plasmodium, and Theileria), the present N-glycan variation is based upon secondary loss of Alg genes, which encode enzymes that add sugars to the N-glycan precursor. An N-glycan precursor with Man5GlcNAc2 is necessary but not sufficient for N-glycan QC, which is predicted by the presence of the UDP-glucose:glucosyltransferase (UGGT) plus calreticulin and/or calnexin. As many parasites lack glucose in their N-glycan precursor, UGGT product may be identified by inhibition of glucosidase II. The presence of an armless calnexin in Toxoplasma suggests secondary loss of N-glycan QC from coccidia. Positive selection for N-glycan sites occurs in secreted proteins of organisms with N-glycan QC and is based upon an increased likelihood of threonine but not serine in the +2 position versus asparagine. In contrast, there appears to be selection against N-glycan length in Plasmodium and N-glycan site density in Toxoplasma. Finally, there is suggestive evidence for N-glycan-dependent ERAD in Trichomonas, which glycosylates and degrades the exogenous reporter mutant carboxypeptidase Y (CPY*).
Keywords: ER-associated degradation; Endoplasmic reticulum; Evolution; N-glycan precursors; Parasites; Quality control of protein folding.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
Quality control of glycoprotein folding and ERAD: the role of N-glycan handling, EDEM1 and OS-9.Histochem Cell Biol. 2017 Feb;147(2):269-284. doi: 10.1007/s00418-016-1513-9. Epub 2016 Nov 1. Histochem Cell Biol. 2017. PMID: 27803995 Review.
-
The evolution of N-glycan-dependent endoplasmic reticulum quality control factors for glycoprotein folding and degradation.Proc Natl Acad Sci U S A. 2007 Jul 10;104(28):11676-81. doi: 10.1073/pnas.0704862104. Epub 2007 Jul 2. Proc Natl Acad Sci U S A. 2007. PMID: 17606910 Free PMC article.
-
Suggestive evidence for Darwinian Selection against asparagine-linked glycans of Plasmodium falciparum and Toxoplasma gondii.Eukaryot Cell. 2010 Feb;9(2):228-41. doi: 10.1128/EC.00197-09. Epub 2009 Sep 25. Eukaryot Cell. 2010. PMID: 19783771 Free PMC article.
-
Role of N-oligosaccharide endoplasmic reticulum processing reactions in glycoprotein folding and degradation.Biochem J. 2000 May 15;348 Pt 1(Pt 1):1-13. Biochem J. 2000. PMID: 10794707 Free PMC article. Review.
-
A sweet code for glycoprotein folding.FEBS Lett. 2015 Nov 14;589(22):3379-87. doi: 10.1016/j.febslet.2015.07.021. Epub 2015 Jul 28. FEBS Lett. 2015. PMID: 26226420 Review.
Cited by
-
Comparative transcriptome analysis of cells from different areas reveals ROS responsive mechanism at sclerotial initiation stage in Morchella importuna.Sci Rep. 2021 May 3;11(1):9418. doi: 10.1038/s41598-021-87784-w. Sci Rep. 2021. PMID: 33941791 Free PMC article.
-
Quality control of glycoprotein folding and ERAD: the role of N-glycan handling, EDEM1 and OS-9.Histochem Cell Biol. 2017 Feb;147(2):269-284. doi: 10.1007/s00418-016-1513-9. Epub 2016 Nov 1. Histochem Cell Biol. 2017. PMID: 27803995 Review.
-
Interdomain conformational flexibility underpins the activity of UGGT, the eukaryotic glycoprotein secretion checkpoint.Proc Natl Acad Sci U S A. 2017 Aug 8;114(32):8544-8549. doi: 10.1073/pnas.1703682114. Epub 2017 Jul 24. Proc Natl Acad Sci U S A. 2017. PMID: 28739903 Free PMC article.
-
N-Glycosylation in Piroplasmids: Diversity within Simplicity.Pathogens. 2021 Jan 8;10(1):50. doi: 10.3390/pathogens10010050. Pathogens. 2021. PMID: 33429911 Free PMC article.
-
Comparative genomics reveals Cyclospora cayetanensis possesses coccidia-like metabolism and invasion components but unique surface antigens.BMC Genomics. 2016 Apr 30;17:316. doi: 10.1186/s12864-016-2632-3. BMC Genomics. 2016. PMID: 27129308 Free PMC article.
References
-
- Morrison HG, McArthur AG, Gillin FD, Aley SB, Adam RD, Olsen GJ, et al. Genomic minimalism in the early diverging intestinal parasite Giardia lamblia. Science. 2007;317:1921–26. - PubMed
-
- Abrahamsen MS, Templeton TJ, Enomoto S, Abrahante JE, Zhu G, Lancto CA, et al. Complete genome sequence of the apicomplexan, Cryptosporidium parvum. Science. 2004;304:441–5. - PubMed
-
- Clark CG, Alsmark UC, Tazreiter M, Saito-Nakano Y, Ali V, Marion S, Weber C, et al. Structure and content of the Entamoeba histolytica genome. Adv Parasitol. 2007;65:51–190. - PubMed
-
- Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H, Bartholomeu DC, et al. The genome of the African trypanosome Trypanosoma brucei. Science. 2005;309:416–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
