

Conservation analysis of the CydX protein yields insights into small protein identification and evolution
- PMID: 25475368
- PMCID: PMC4325964
- DOI: 10.1186/1471-2164-15-946
Conservation analysis of the CydX protein yields insights into small protein identification and evolution
Abstract
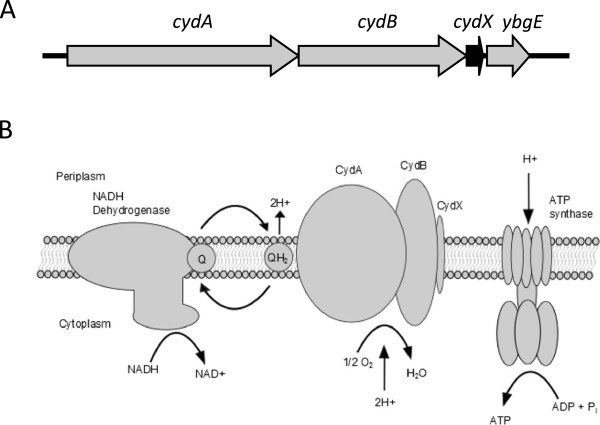
Background: The reliable identification of proteins containing 50 or fewer amino acids is difficult due to the limited information content in short sequences. The 37 amino acid CydX protein in Escherichia coli is a member of the cytochrome bd oxidase complex, an enzyme found throughout Eubacteria. To investigate the extent of CydX conservation and prevalence and evaluate different methods of small protein homologue identification, we surveyed 1095 Eubacteria species for the presence of the small protein.
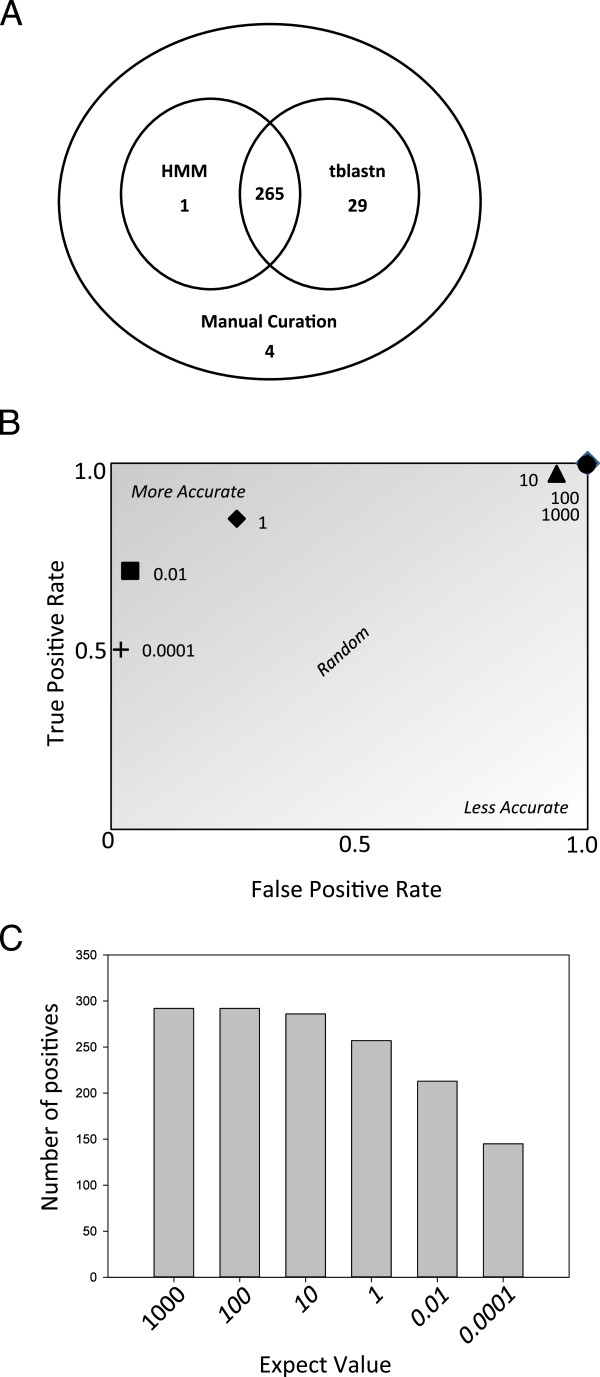
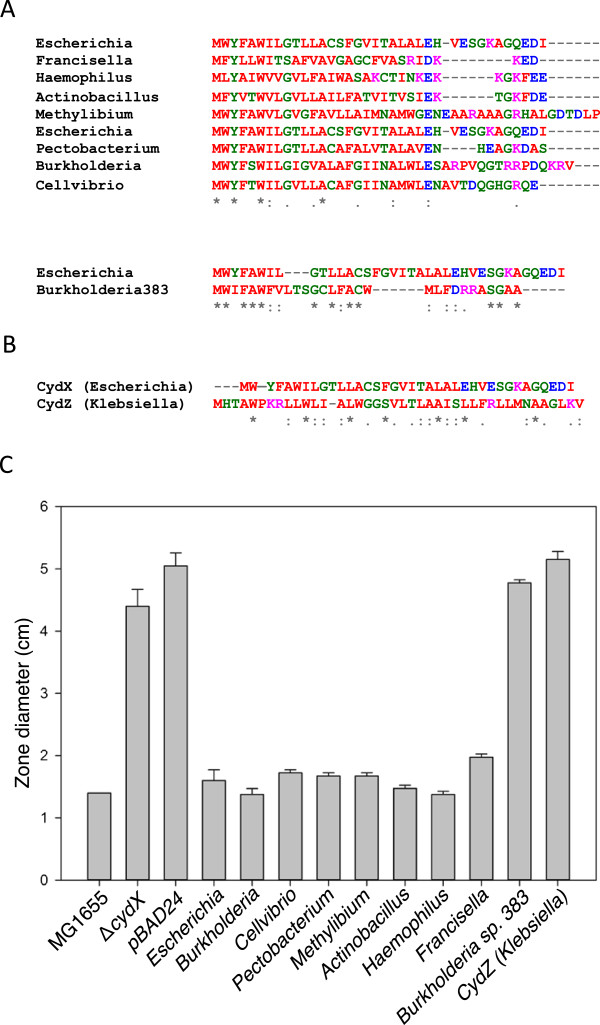
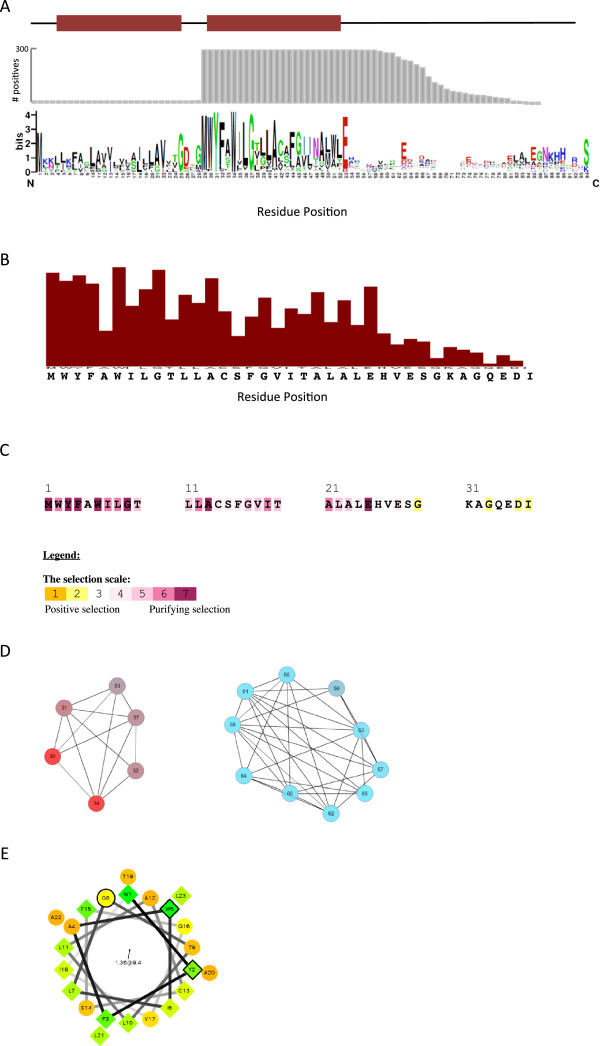
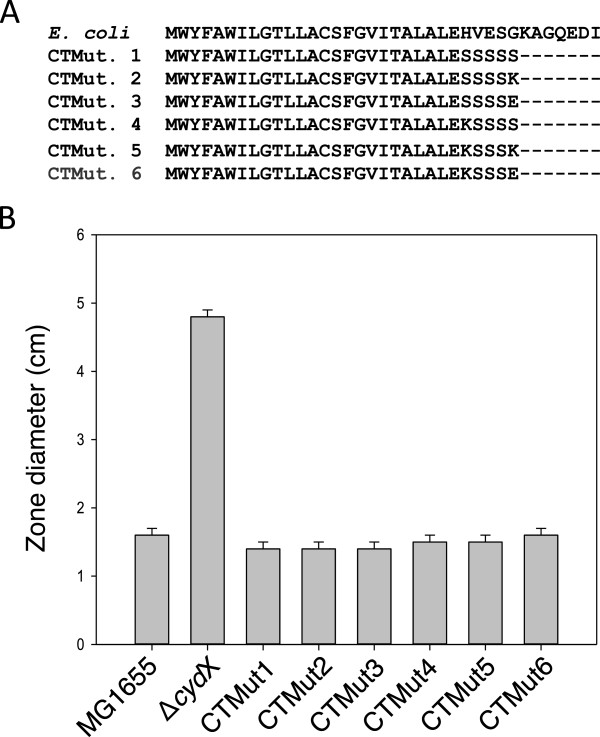
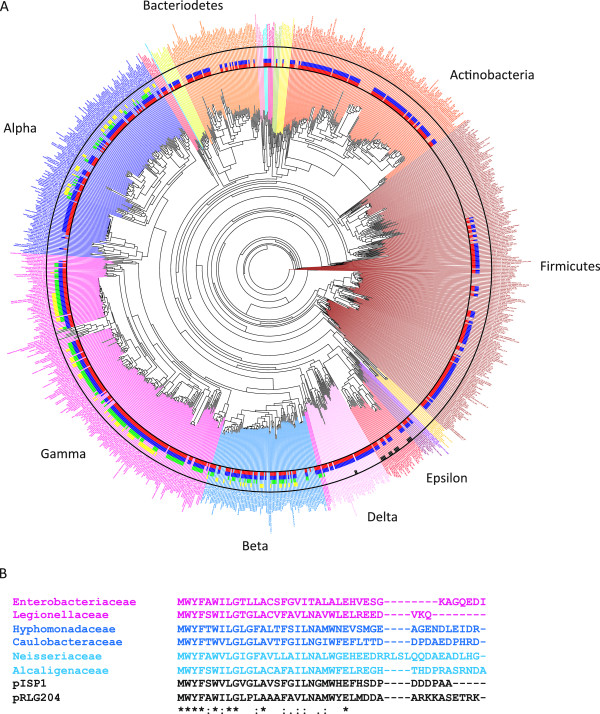
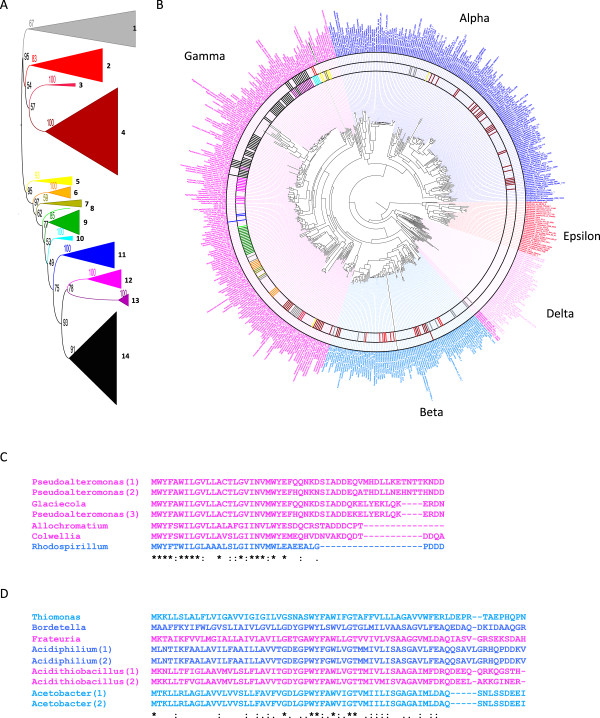
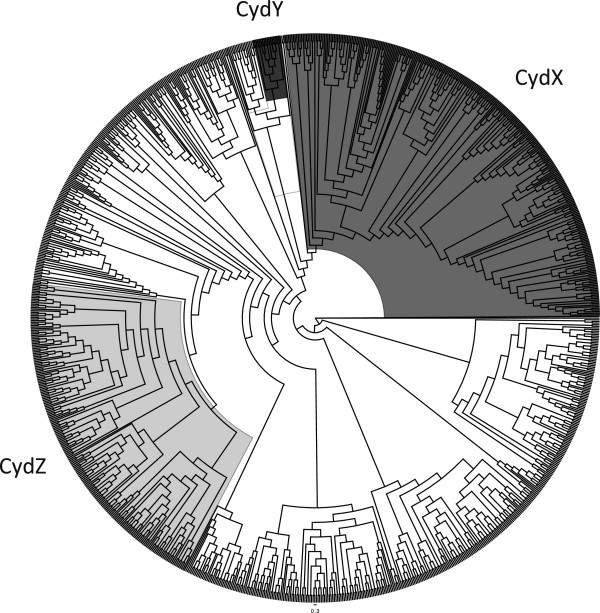
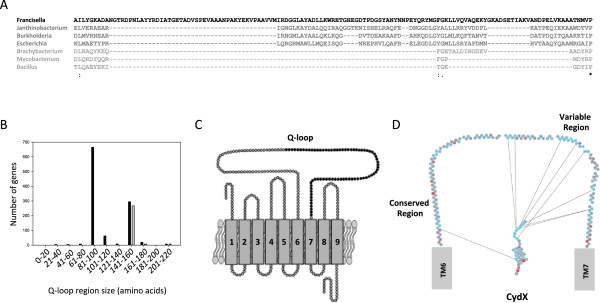
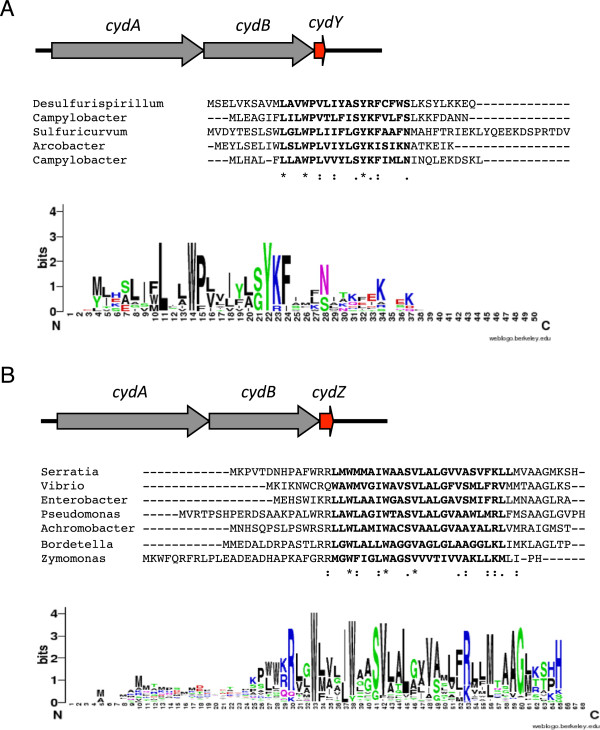
Results: Over 300 homologues were identified, including 80 unannotated genes. The ability of both closely-related and divergent homologues to complement the E. coli ΔcydX mutant supports our identification techniques, and suggests that CydX homologues retain similar function among divergent species. However, sequence analysis of these proteins shows a great degree of variability, with only a few highly-conserved residues. An analysis of the co-variation between CydX homologues and their corresponding cydA and cydB genes shows a close synteny of the small protein with the CydA long Q-loop. Phylogenetic analysis suggests that the cydABX operon has undergone horizontal gene transfer, although the cydX gene likely evolved in a progenitor of the Alpha, Beta, and Gammaproteobacteria. Further investigation of cydAB operons identified two additional conserved hypothetical small proteins: CydY encoded in CydAQlong operons that lack cydX, and CydZ encoded in more than 150 CydAQshort operons.
Conclusions: This study provides a systematic analysis of bioinformatics techniques required for the unique challenges present in small protein identification and phylogenetic analyses. These results elucidate the prevalence of CydX throughout the Proteobacteria, provide insight into the selection pressure and sequence requirements for CydX function, and suggest a potential functional interaction between the small protein and the CydA Q-loop, an enigmatic domain of the cytochrome bd oxidase complex. Finally, these results identify other conserved small proteins encoded in cytochrome bd oxidase operons, suggesting that small protein subunits may be a more common component of these enzymes than previously thought.
Figures
Comment in
-
The small protein floodgates are opening; now the functional analysis begins.BMC Biol. 2014 Dec 5;12:96. doi: 10.1186/s12915-014-0096-y. BMC Biol. 2014. PMID: 25475548 Free PMC article.
Similar articles
-
The Escherichia coli CydX protein is a member of the CydAB cytochrome bd oxidase complex and is required for cytochrome bd oxidase activity.J Bacteriol. 2013 Aug;195(16):3640-50. doi: 10.1128/JB.00324-13. Epub 2013 Jun 7. J Bacteriol. 2013. PMID: 23749980 Free PMC article.
-
Investigation of amino acid specificity in the CydX small protein shows sequence plasticity at the functional level.PLoS One. 2018 Jun 18;13(6):e0198699. doi: 10.1371/journal.pone.0198699. eCollection 2018. PLoS One. 2018. PMID: 29912917 Free PMC article.
-
Subunit CydX of Escherichia coli cytochrome bd ubiquinol oxidase is essential for assembly and stability of the di-heme active site.FEBS Lett. 2014 May 2;588(9):1537-41. doi: 10.1016/j.febslet.2014.03.036. Epub 2014 Mar 26. FEBS Lett. 2014. PMID: 24681096
-
Oxygen as Acceptor.EcoSal Plus. 2015;6(2):10.1128/ecosalplus.ESP-0012-2015. doi: 10.1128/ecosalplus.ESP-0012-2015. EcoSal Plus. 2015. PMID: 26734697 Free PMC article. Review.
-
Cytochrome bd terminal oxidase.Biochim Biophys Acta. 1997 Aug 22;1321(2):107-27. doi: 10.1016/s0005-2728(97)00046-7. Biochim Biophys Acta. 1997. PMID: 9332500 Review. No abstract available.
Cited by
-
Recent Advances in Structural Studies of Cytochrome bd and Its Potential Application as a Drug Target.Int J Mol Sci. 2022 Mar 15;23(6):3166. doi: 10.3390/ijms23063166. Int J Mol Sci. 2022. PMID: 35328590 Free PMC article. Review.
-
Escherichia coli Small Proteome.EcoSal Plus. 2020 May;9(1):10.1128/ecosalplus.ESP-0031-2019. doi: 10.1128/ecosalplus.ESP-0031-2019. EcoSal Plus. 2020. PMID: 32385980 Free PMC article. Review.
-
Uncovering the small proteome of Methanosarcina mazei using Ribo-seq and peptidomics under different nitrogen conditions.Nat Commun. 2024 Oct 6;15(1):8659. doi: 10.1038/s41467-024-53008-8. Nat Commun. 2024. PMID: 39370430 Free PMC article.
-
PIRT-Seq: a high-resolution whole-genome assay to identify protein-coding genes.Nucleic Acids Res. 2025 Aug 11;53(15):gkaf774. doi: 10.1093/nar/gkaf774. Nucleic Acids Res. 2025. PMID: 40808296 Free PMC article.
-
Alternative ORFs and small ORFs: shedding light on the dark proteome.Nucleic Acids Res. 2020 Feb 20;48(3):1029-1042. doi: 10.1093/nar/gkz734. Nucleic Acids Res. 2020. PMID: 31504789 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
