

An integrated biological approach to guide the development of metal-chelating inhibitors of influenza virus PA endonuclease
- PMID: 25477342
- PMCID: PMC11037440
- DOI: 10.1124/mol.114.095588
An integrated biological approach to guide the development of metal-chelating inhibitors of influenza virus PA endonuclease
Abstract
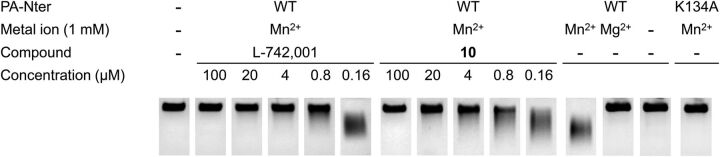
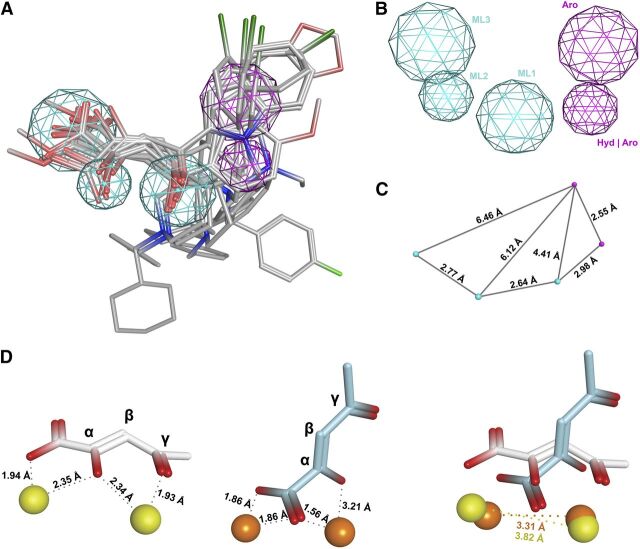
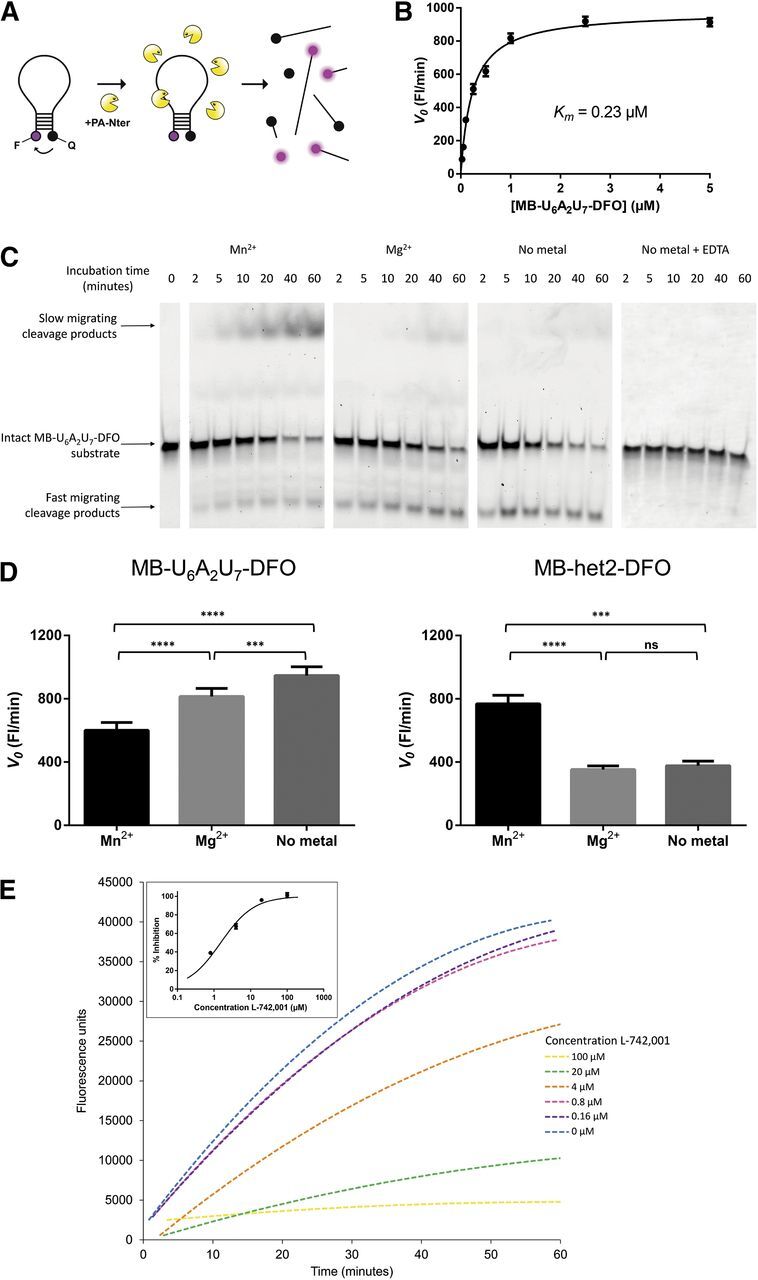
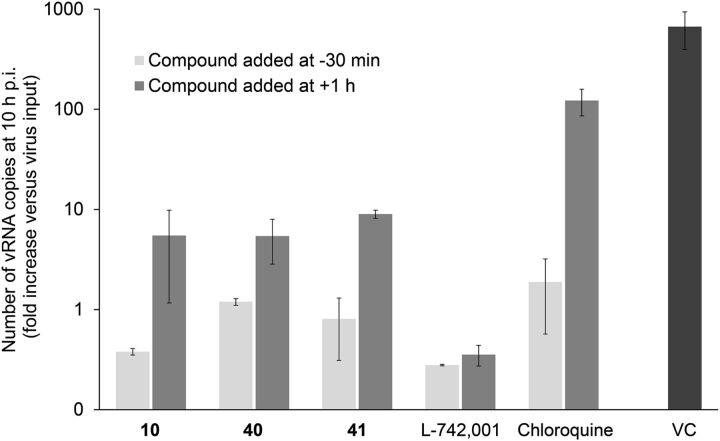
The influenza virus PA endonuclease, which cleaves capped cellular pre-mRNAs to prime viral mRNA synthesis, is a promising target for novel anti-influenza virus therapeutics. The catalytic center of this enzyme resides in the N-terminal part of PA (PA-Nter) and contains two (or possibly one or three) Mg(2+) or Mn(2+) ions, which are critical for its catalytic function. There is great interest in PA inhibitors that are optimally designed to occupy the active site and chelate the metal ions. We focused here on a series of β-diketo acid (DKA) and DKA-bioisosteric compounds containing different scaffolds, and determined their structure-activity relationship in an enzymatic assay with PA-Nter, in order to build a three-dimensional pharmacophore model. In addition, we developed a molecular beacon (MB)-based PA-Nter assay that enabled us to compare the inhibition of Mn(2+) versus Mg(2+), the latter probably being the biologically relevant cofactor. This real-time MB assay allowed us to measure the enzyme kinetics of PA-Nter or perform high-throughput screening. Several DKA derivatives were found to cause strong inhibition of PA-Nter, with IC50 values comparable to that of the prototype L-742,001 (i.e., below 2 μM). Among the different compounds tested, L-742,001 appeared unique in having equal activity against either Mg(2+) or Mn(2+). Three compounds ( 10: , with a pyrrole scaffold, and 40: and 41: , with an indole scaffold) exhibited moderate antiviral activity in cell culture (EC99 values 64-95 μM) and were proven to affect viral RNA synthesis. Our approach of integrating complementary enzymatic, cellular, and mechanistic assays should guide ongoing development of improved influenza virus PA inhibitors.
Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics.
Figures
References
-
- Bacchi A, Biemmi M, Carcelli M, Carta F, Compari C, Fisicaro E, Rogolino D, Sechi M, Sippel M, Sotriffer CA, et al. (2008) From ligand to complexes. Part 2. Remarks on human immunodeficiency virus type 1 integrase inhibition by beta-diketo acid metal complexes. J Med Chem 51:7253–7264. - PubMed
-
- Bacchi A, Carcelli M, Compari C, Fisicaro E, Pala N, Rispoli G, Rogolino D, Sanchez TW, Sechi M, Sinisi V, et al. (2011) Investigating the role of metal chelation in HIV-1 integrase strand transfer inhibitors. J Med Chem 54:8407–8420. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
