

Neural Differentiation Tracks Improved Recall of Competing Memories Following Interleaved Study and Retrieval Practice
- PMID: 25477369
- PMCID: PMC4585527
- DOI: 10.1093/cercor/bhu284
Neural Differentiation Tracks Improved Recall of Competing Memories Following Interleaved Study and Retrieval Practice
Abstract
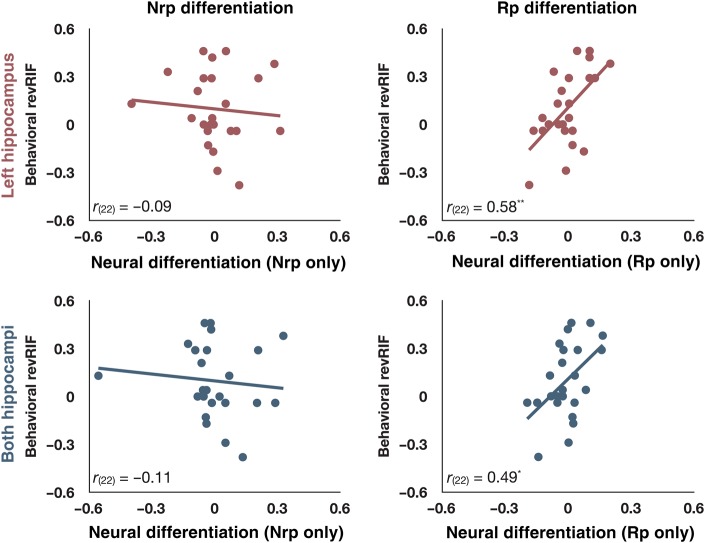
Selective retrieval of overlapping memories can generate competition. How does the brain adaptively resolve this competition? One possibility is that competing memories are inhibited; in support of this view, numerous studies have found that selective retrieval leads to forgetting of memories that are related to the just-retrieved memory. However, this retrieval-induced forgetting (RIF) effect can be eliminated or even reversed if participants are given opportunities to restudy the materials between retrieval attempts. Here, we outline an explanation for such a reversal, rooted in a neural network model of RIF that predicts representational differentiation when restudy is interleaved with selective retrieval. To test this hypothesis, we measured changes in pattern similarity of the BOLD fMRI signal elicited by related memories after undergoing interleaved competitive retrieval and restudy. Reduced pattern similarity within the hippocampus positively correlated with retrieval-induced facilitation of competing memories. This result is consistent with an adaptive differentiation process that allows individuals to learn to distinguish between once-confusable memories.
Keywords: differentiation; hippocampus; memory; neural network model; pattern similarity; retrieval-induced forgetting.
© The Author 2014. Published by Oxford University Press. All rights reserved. For Permissions, please e-mail: journals.permissions@oup.com.
Figures


Similar articles
-
A neural network model of retrieval-induced forgetting.Psychol Rev. 2007 Oct;114(4):887-953. doi: 10.1037/0033-295X.114.4.887. Psychol Rev. 2007. PMID: 17907868
-
Retrieval from episodic memory: neural mechanisms of interference resolution.J Cogn Neurosci. 2009 Mar;21(3):538-49. doi: 10.1162/jocn.2009.21043. J Cogn Neurosci. 2009. PMID: 18564040
-
Neural Differentiation of Incorrectly Predicted Memories.J Neurosci. 2017 Feb 22;37(8):2022-2031. doi: 10.1523/JNEUROSCI.3272-16.2017. Epub 2017 Jan 23. J Neurosci. 2017. PMID: 28115478 Free PMC article.
-
Top-down and bottom-up attention to memory: a hypothesis (AtoM) on the role of the posterior parietal cortex in memory retrieval.Neuropsychologia. 2008;46(7):1828-51. doi: 10.1016/j.neuropsychologia.2008.03.022. Epub 2008 Apr 8. Neuropsychologia. 2008. PMID: 18471837 Review.
-
Towards a cognitive and neurobiological model of motivated forgetting.Nebr Symp Motiv. 2012;58:53-120. doi: 10.1007/978-1-4614-1195-6_3. Nebr Symp Motiv. 2012. PMID: 22303764 Review.
Cited by
-
Gaze-pattern similarity at encoding may interfere with future memory.Sci Rep. 2021 Apr 8;11(1):7697. doi: 10.1038/s41598-021-87258-z. Sci Rep. 2021. PMID: 33833314 Free PMC article.
-
Partially overlapping spatial environments trigger reinstatement in hippocampus and schema representations in prefrontal cortex.Nat Commun. 2021 Oct 28;12(1):6231. doi: 10.1038/s41467-021-26560-w. Nat Commun. 2021. PMID: 34711830 Free PMC article.
-
Rapid neural reorganization during retrieval practice predicts subsequent long-term retention and false memory.Nat Hum Behav. 2022 Jan;6(1):134-145. doi: 10.1038/s41562-021-01188-4. Epub 2021 Oct 7. Nat Hum Behav. 2022. PMID: 34621051
-
Differentiation and Integration of Competing Memories: A Neural Network Model.bioRxiv [Preprint]. 2024 Jun 25:2023.04.02.535239. doi: 10.1101/2023.04.02.535239. bioRxiv. 2024. Update in: Elife. 2024 Sep 25;12:RP88608. doi: 10.7554/eLife.88608. PMID: 37066178 Free PMC article. Updated. Preprint.
-
Adaptive Memory Distortions Are Predicted by Feature Representations in Parietal Cortex.J Neurosci. 2021 Mar 31;41(13):3014-3024. doi: 10.1523/JNEUROSCI.2875-20.2021. Epub 2021 Feb 22. J Neurosci. 2021. PMID: 33619210 Free PMC article.
References
-
- Abel M, Bäuml K-HT. 2012. Retrieval-induced forgetting, delay, and sleep. Memory. 20:420–428. - PubMed
-
- Anderson MC. 2003. Rethinking interference theory: executive control and the mechanisms of forgetting. J Mem Lang. 49:415–445.
-
- Anderson MC, Bjork RA, Bjork EL. 1994. Remembering can cause forgetting: retrieval dynamics in long-term memory. J Exp Psychol Learn. 20:1063–1087. - PubMed
-
- Anderson MC, McCulloch KC. 1999. Integration as a general boundary condition on retrieval-induced forgetting. J Experim Psychol. 25:608–629.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
