

Chromatin decondensation is sufficient to alter nuclear organization in embryonic stem cells
- PMID: 25477464
- PMCID: PMC6529354
- DOI: 10.1126/science.1259587
Chromatin decondensation is sufficient to alter nuclear organization in embryonic stem cells
Abstract
During differentiation, thousands of genes are repositioned toward or away from the nuclear envelope. These movements correlate with changes in transcription and replication timing. Using synthetic (TALE) transcription factors, we found that transcriptional activation of endogenous genes by a viral trans-activator is sufficient to induce gene repositioning toward the nuclear interior in embryonic stem cells. However, gene relocation was also induced by recruitment of an acidic peptide that decondenses chromatin without affecting transcription, indicating that nuclear reorganization is driven by chromatin remodeling rather than transcription. We identified an epigenetic inheritance of chromatin decondensation that maintained central nuclear positioning through mitosis even after the TALE transcription factor was lost. Our results also demonstrate that transcriptional activation, but not chromatin decondensation, is sufficient to change replication timing.
Copyright © 2014, American Association for the Advancement of Science.
Figures
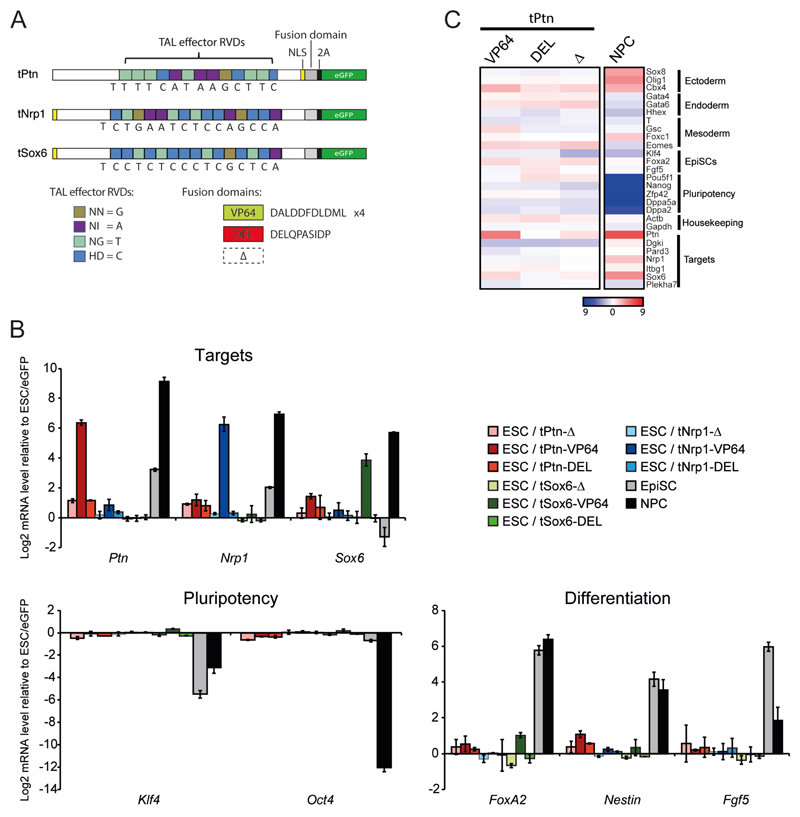
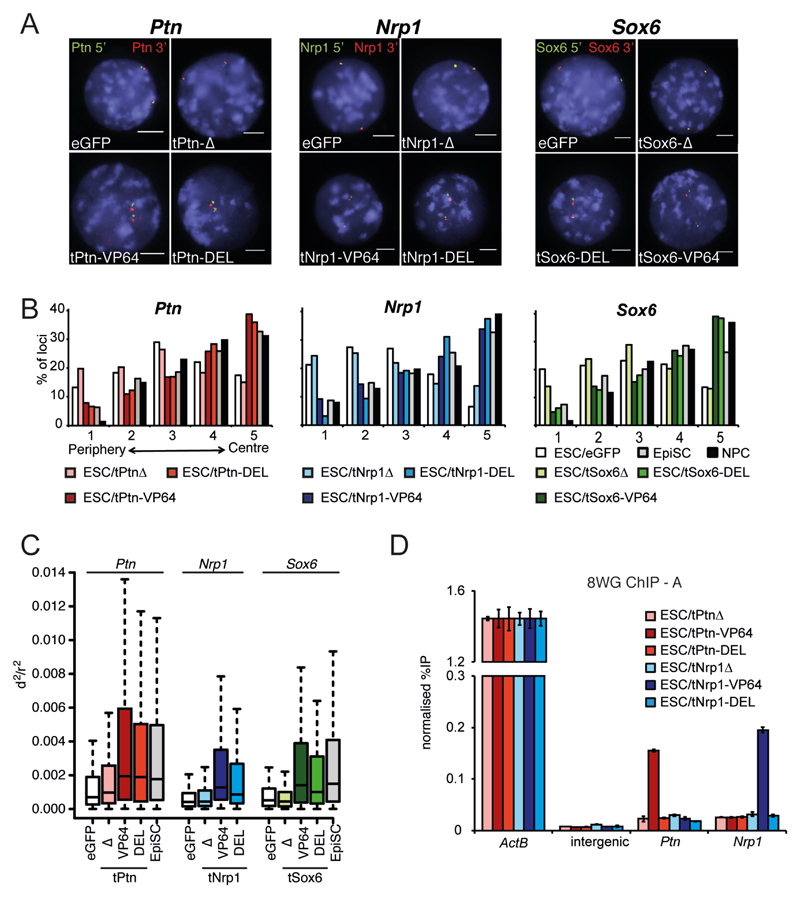
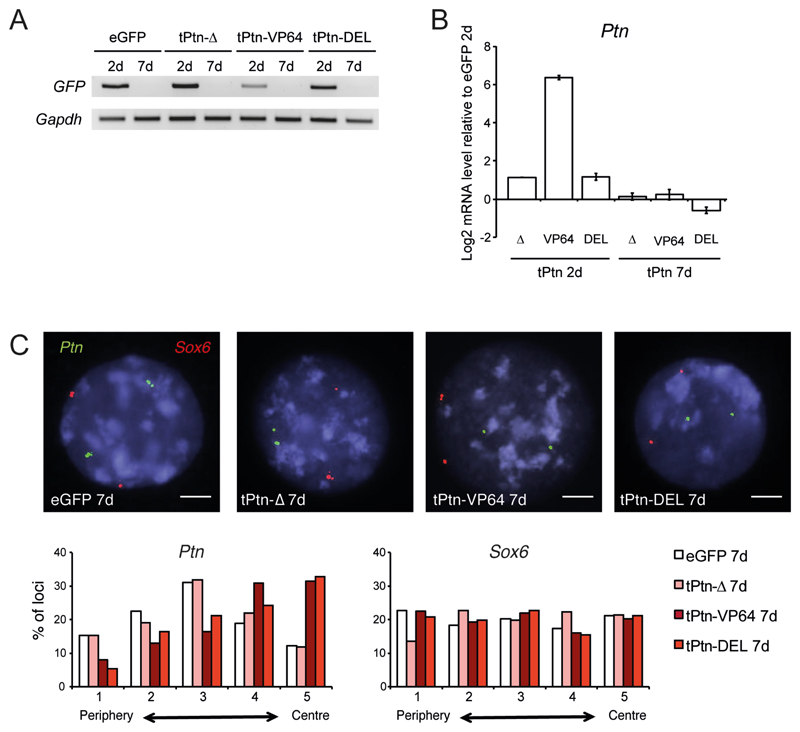
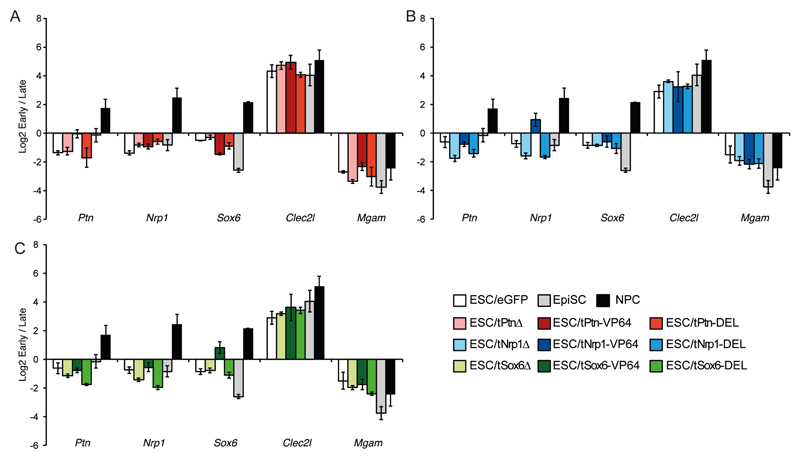
Comment in
-
Chromatin. Drivers of nuclear organization.Nat Rev Mol Cell Biol. 2015 Feb;16(2):67. doi: 10.1038/nrm3938. Nat Rev Mol Cell Biol. 2015. PMID: 25604199
References
-
- Meister P, Taddei A. Building silent compartments at the nuclear periphery: a recurrent theme. Current opinion in genetics & development. 2013;23:96–103. - PubMed
-
- Akhtar A, Gasser SM. The nuclear envelope and transcriptional control. Nature reviews. Genetics. 2007;8:507–517. - PubMed
-
- Guelen L, Pagie L, Brasset E, Meuleman W, Faza MB, Talhout W, et al. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature. 2008;453:948–951. - PubMed
-
- Reddy KL, Zullo JM, Bertolino E, Singh H. Transcriptional repression mediated by repositioning of genes to the nuclear lamina. Nature. 2008;452:243–247. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
