

Cardiac repair in a porcine model of acute myocardial infarction with human induced pluripotent stem cell-derived cardiovascular cells
- PMID: 25479750
- PMCID: PMC4275050
- DOI: 10.1016/j.stem.2014.11.009
Cardiac repair in a porcine model of acute myocardial infarction with human induced pluripotent stem cell-derived cardiovascular cells
Erratum in
- Cell Stem Cell. 2015 Jan 8;16(1):102
Abstract
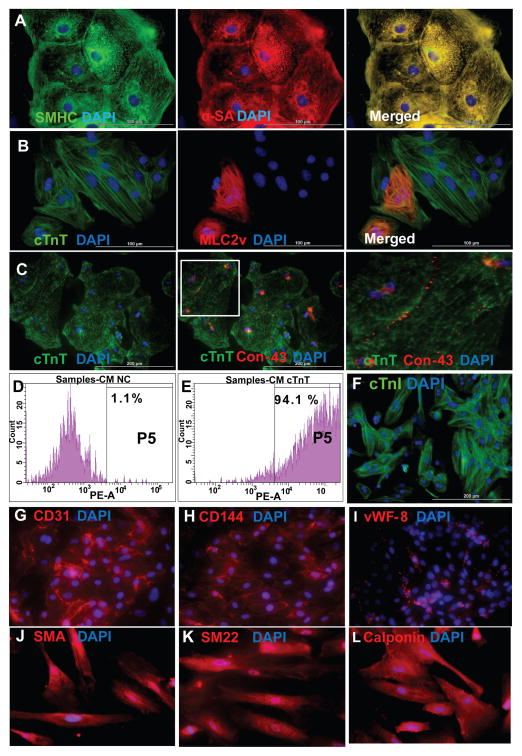
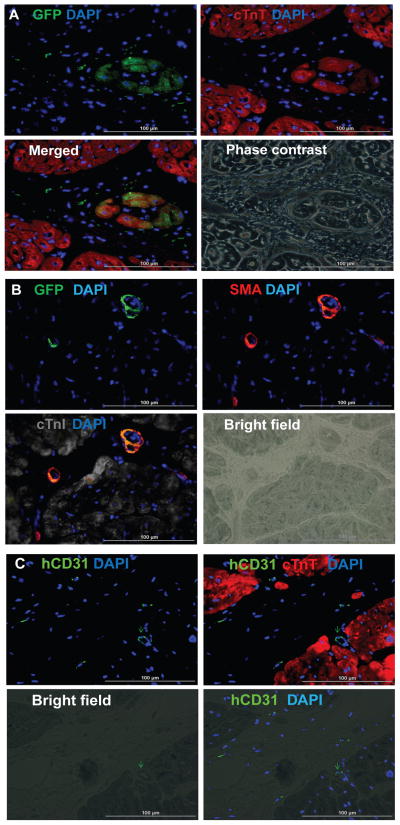
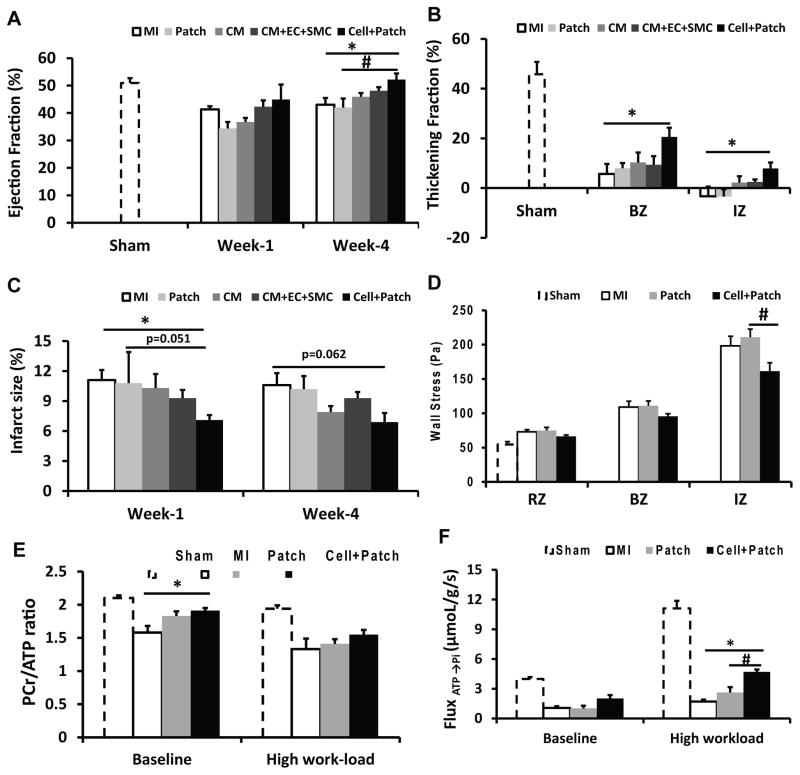
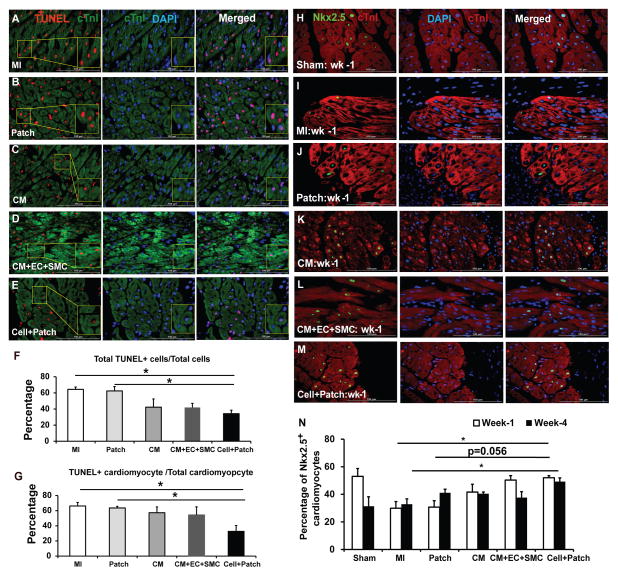
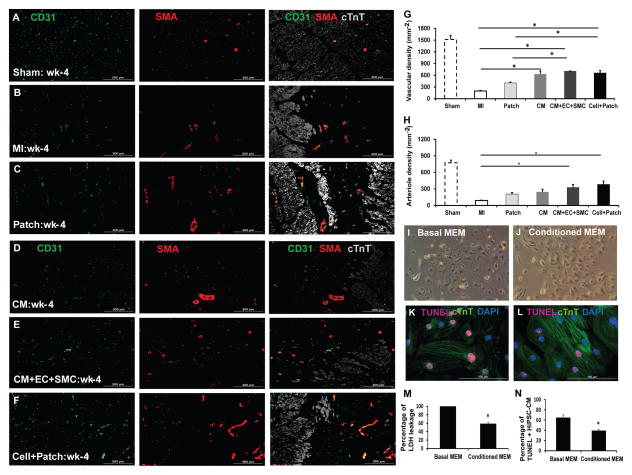
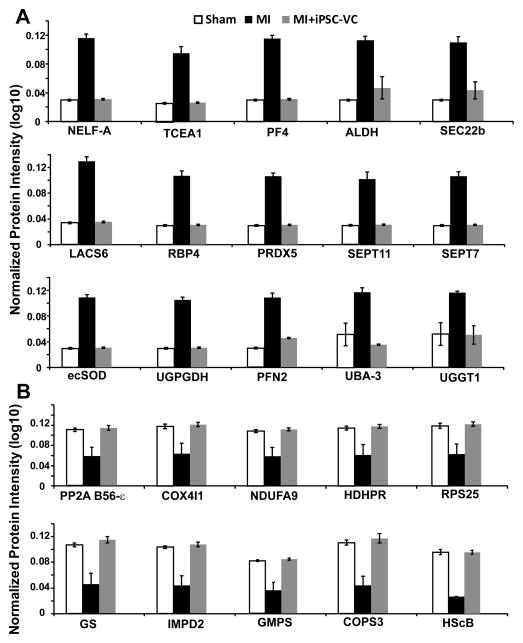
Human induced pluripotent stem cells (hiPSCs) hold promise for myocardial repair following injury, but preclinical studies in large animal models are required to determine optimal cell preparation and delivery strategies to maximize functional benefits and to evaluate safety. Here, we utilized a porcine model of acute myocardial infarction (MI) to investigate the functional impact of intramyocardial transplantation of hiPSC-derived cardiomyocytes, endothelial cells, and smooth muscle cells, in combination with a 3D fibrin patch loaded with insulin growth factor (IGF)-encapsulated microspheres. hiPSC-derived cardiomyocytes integrated into host myocardium and generated organized sarcomeric structures, and endothelial and smooth muscle cells contributed to host vasculature. Trilineage cell transplantation significantly improved left ventricular function, myocardial metabolism, and arteriole density, while reducing infarct size, ventricular wall stress, and apoptosis without inducing ventricular arrhythmias. These findings in a large animal MI model highlight the potential of utilizing hiPSC-derived cells for cardiac repair.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
Comment in
-
Patching up broken hearts: cardiac cell therapy gets a bioengineered boost.Cell Stem Cell. 2014 Dec 4;15(6):671-3. doi: 10.1016/j.stem.2014.11.008. Cell Stem Cell. 2014. PMID: 25479741 Free PMC article.
References
-
- Brutsaert DL. Cardiac endothelial-myocardial signaling: its role in cardiac growth, contractile performance, and rhythmicity. Physiological reviews. 2003;83:59–115. - PubMed
-
- Caspi O, Huber I, Kehat I, Habib M, Arbel G, Gepstein A, Yankelson L, Aronson D, Beyar R, Gepstein L. Transplantation of human embryonic stem cell-derived cardiomyocytes improves myocardial performance in infarcted rat hearts. Journal of the American College of Cardiology. 2007;50:1884–1893. - PubMed
-
- Chen D, Dorling A. Critical roles for thrombin in acute and chronic inflammation. Journal of thrombosis and haemostasis: JTH. 2009;7(Suppl 1):122–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
