

Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery
- PMID: 25480295
- PMCID: PMC4258230
- DOI: 10.1016/j.cell.2014.11.017
Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery
Abstract
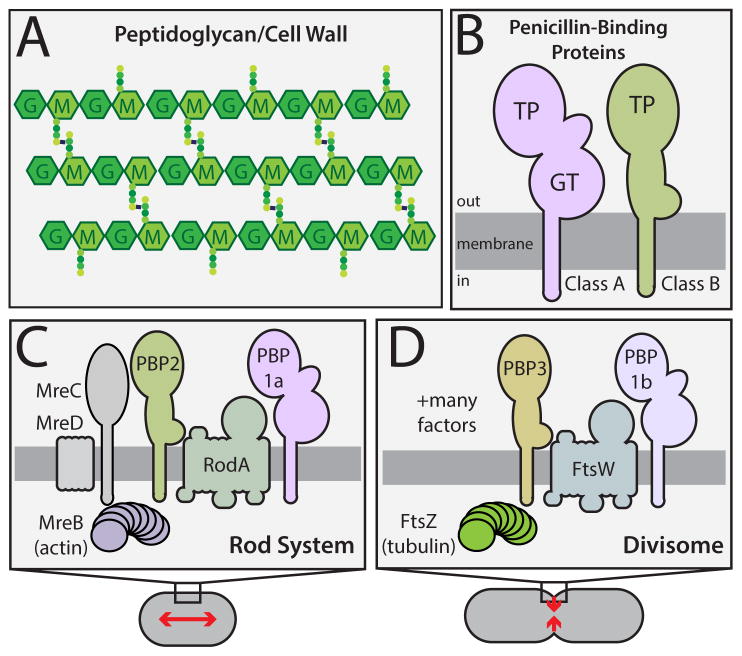
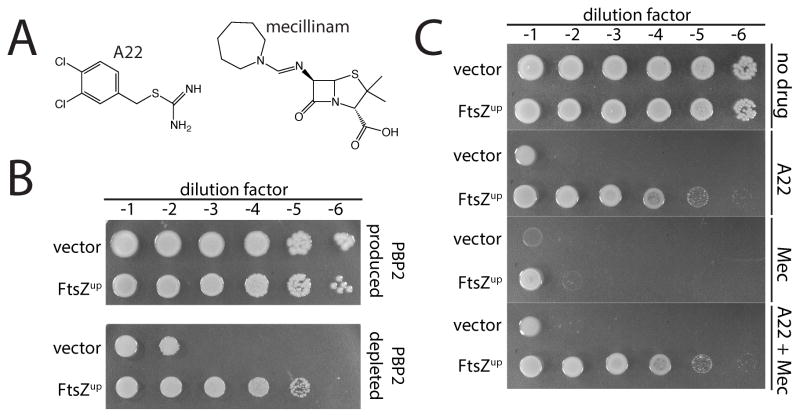
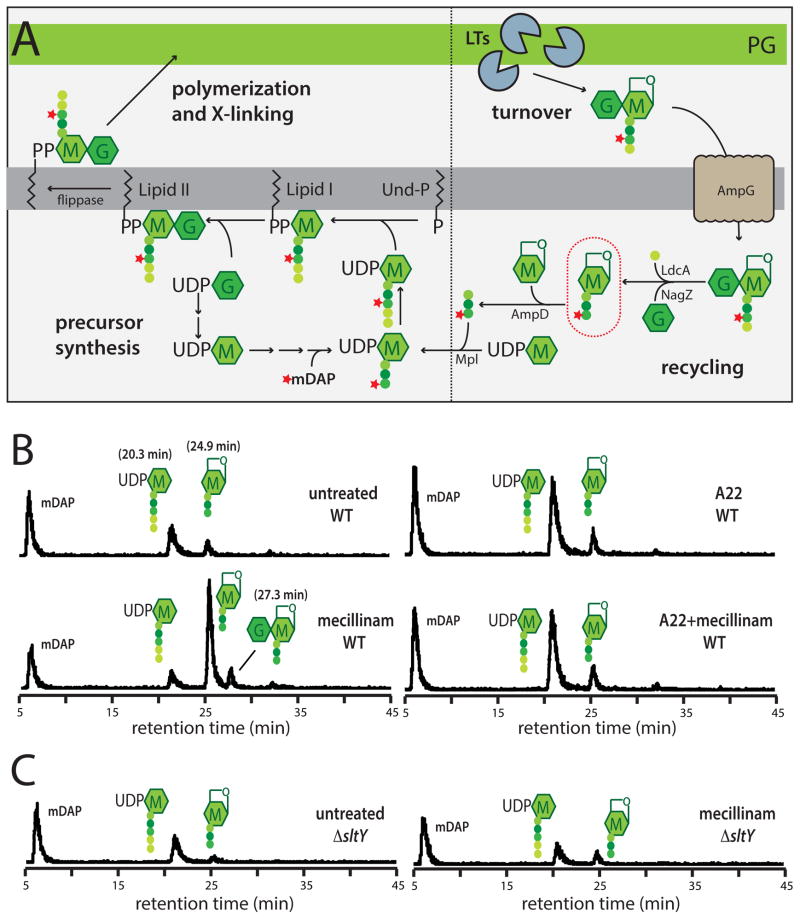
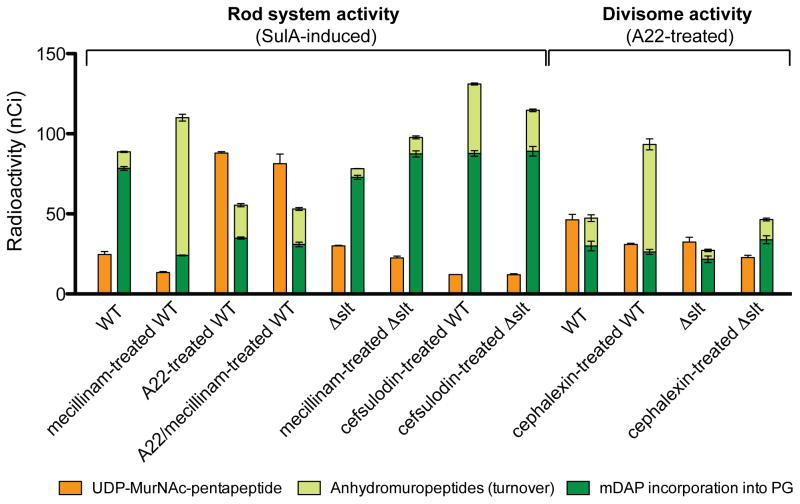
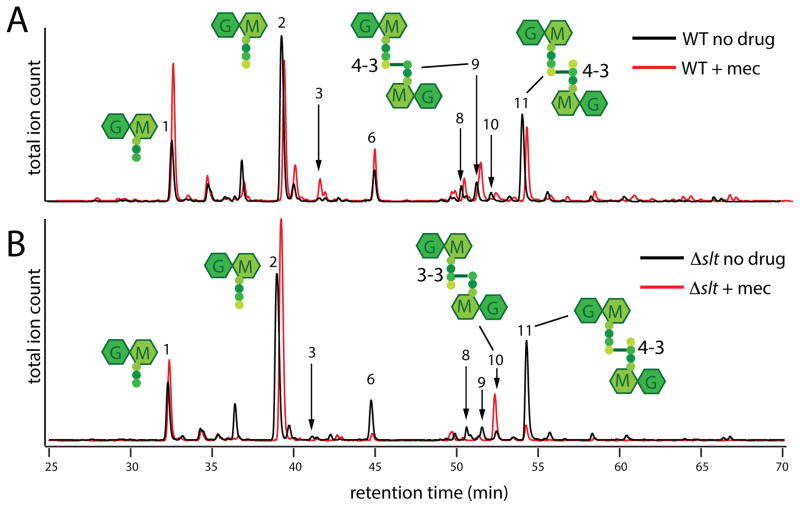
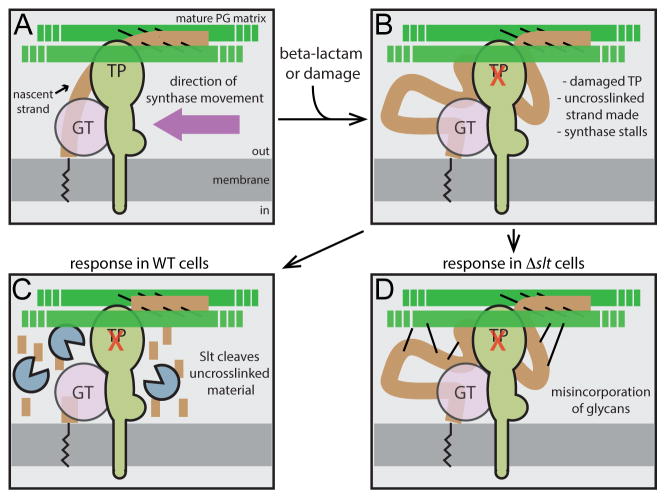
Penicillin and related beta-lactams comprise one of our oldest and most widely used antibiotic therapies. These drugs have long been known to target enzymes called penicillin-binding proteins (PBPs) that build the bacterial cell wall. Investigating the downstream consequences of target inhibition and how they contribute to the lethal action of these important drugs, we demonstrate that beta-lactams do more than just inhibit the PBPs as is commonly believed. Rather, they induce a toxic malfunctioning of their target biosynthetic machinery involving a futile cycle of cell wall synthesis and degradation, thereby depleting cellular resources and bolstering their killing activity. Characterization of this mode of action additionally revealed a quality control function for enzymes that cleave bonds in the cell wall matrix. The results thus provide insight into the mechanism of cell wall assembly and suggest how best to interfere with the process for future antibiotic development.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures

Comment in
-
Antimicrobials: New tricks for old drugs.Nat Rev Microbiol. 2015 Feb;13(2):68. doi: 10.1038/nrmicro3421. Epub 2014 Dec 22. Nat Rev Microbiol. 2015. PMID: 25534806 No abstract available.
References
-
- Banzhaf M, van den Berg van Saparoea B, Terrak M, Fraipont C, Egan A, Philippe J, Zapun A, Breukink E, Nguyen-Distèche M, den Blaauwen T, et al. Cooperativity of peptidoglycan synthases active in bacterial cell elongation. Mol Microbiol. 2012;85:179–194. - PubMed
-
- Beachey EH, Keck W, de Pedro MA, Schwarz U. Exoenzymatic activity of transglycosylase isolated from Escherichia coli. Eur J Biochem. 1981;116:355–358. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
