

Review
doi: 10.1007/978-1-4939-2095-2_10.
A systems biology approach to iron metabolism
Affiliations
- PMID: 25480643
- PMCID: PMC4464783
- DOI: 10.1007/978-1-4939-2095-2_10
Item in Clipboard
Review
A systems biology approach to iron metabolism
Adv Exp Med Biol.
2014.
Abstract
Iron is critical to the survival of almost all living organisms. However, inappropriately low or high levels of iron are detrimental and contribute to a wide range of diseases. Recent advances in the study of iron metabolism have revealed multiple intricate pathways that are essential to the maintenance of iron homeostasis. Further, iron regulation involves processes at several scales, ranging from the subcellular to the organismal. This complexity makes a systems biology approach crucial, with its enabling technology of computational models based on a mathematical description of regulatory systems. Systems biology may represent a new strategy for understanding imbalances in iron metabolism and their underlying causes.
Figures
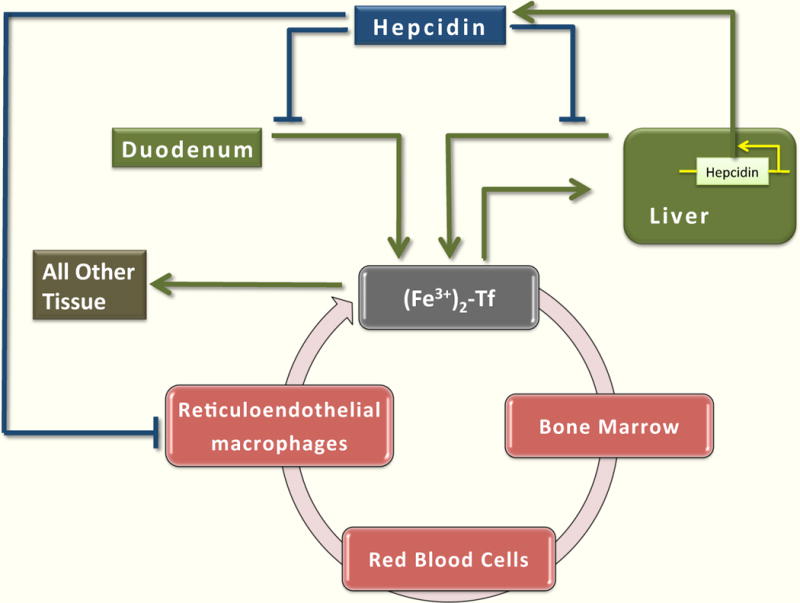
Inorganic, nonheme iron is absorbed by duodenal enterocytes. In the plasma, iron circulates bound to transferrin (Tf). The principal consumer of iron is the erythroid bone marrow, and most of that iron comes from internal recycling by tissue macrophages, predominantly splenic macrophages. Liver is the major storage site of iron. Iron entering the liver is stored in ferritin and can be mobilized when required by the body. Some iron is incorporated in other tissues. Hepcidin regulates systemic iron homeostasis by inhibiting iron release from duodenal enterocytes, macrophages, and hepatocytes.
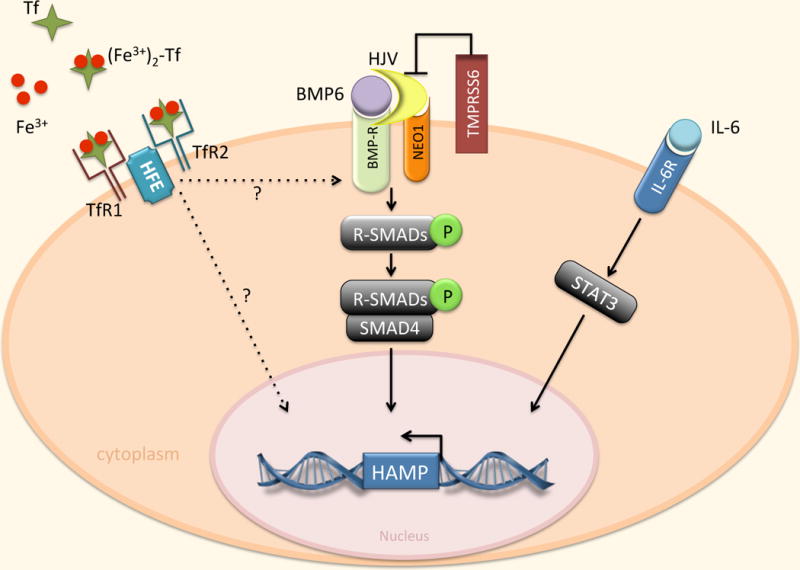
Regulation of hepcidin by BMP/SMAD and IL-6/STAT3 pathways. Expression of hepcidin in the liver is mainly affected by transcriptional mechanisms mediated by the BMP family, primarily BMP6. BMP binds to its receptor (BMPR) in conjunction with the co-receptor HJV. This interaction induces the phosphorylation of R-SMAD proteins which interact with the common mediator SMAD4, bind specific sequences in the hepcidin promoter, and trigger hepcidin gene (HAMP) transcription. NEO1 may enhance BMP signaling by interacting with HJV. TMPRSS6 negatively regulates hepcidin by cleaving HJV. Hepcidin expression is also induced by IL-6 through activation of STAT3. STAT3 binds to specific sequences in the HAMP promoter. TfR2 and HFE are also involved in hepcidin activation through mechanisms that are incompletely defined.
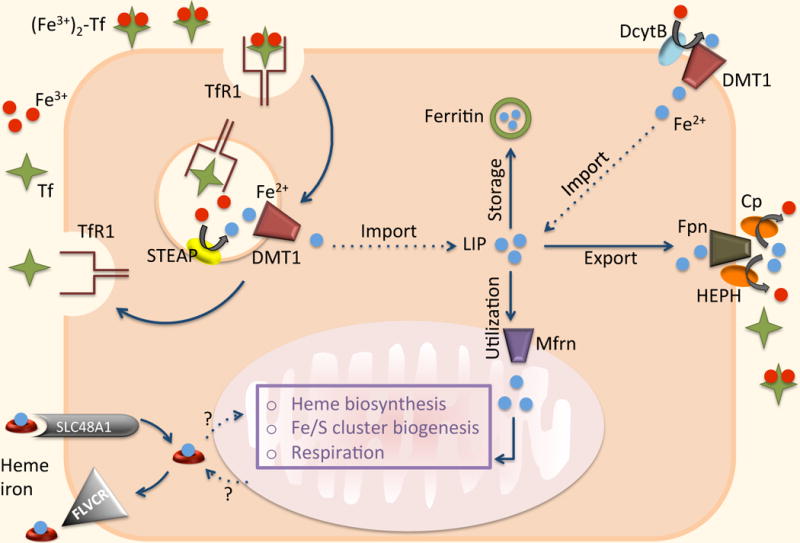
Cells acquire iron predominantly via TfR1. Ferric iron is reduced to Fe2+ by the transmembrane family of metalloreductases (STEAP). DMT1 then facilitates the transport of ferrous iron from the endosomes into the cytoplasm. In some cells, e.g. enterocytes, DMT1 participates in the transport of extracellular iron. DcytB reduces nonheme iron to Fe2+ before it is transported through the cellular membrane. Following egress from the endosome, iron enters the so-called “labile iron pool” (LIP). Ferroportin (Fpn) is believed to be the only ferrous iron exporter. It has been suggested that dietary heme iron can enter through SLC48A1 and be exported via FLVCR. A mechanism by which heme is moved in and out of mitochondria (the major site of iron utilization) is poorly understood. Ferrous iron is imported into mitochondria for incorporation into bioactive heme by the SLC transporter mitoferrin (Mfrn). Iron that is not exported or utilized is stored in ferritin (Ft).
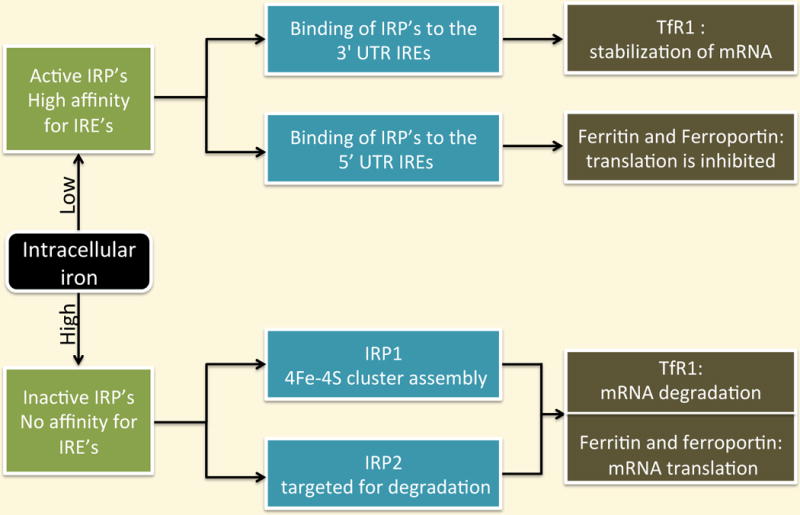
Intracellular iron regulation
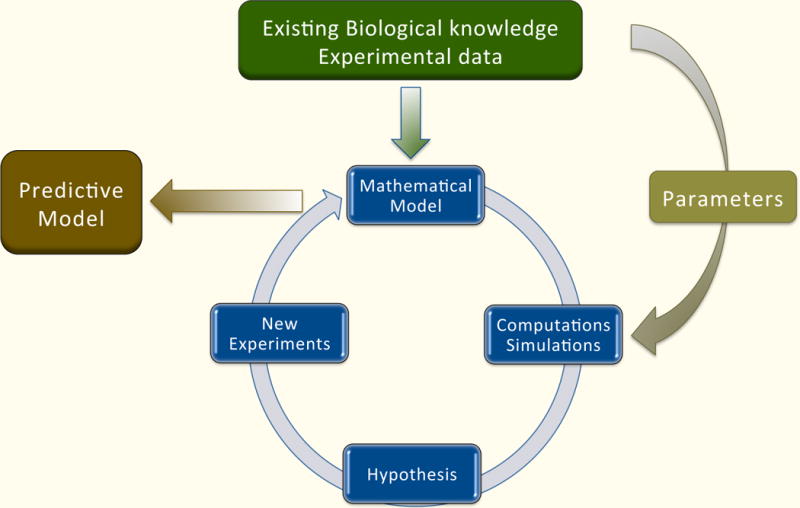
The starting point is a formulation of the problem and specific questions that the model will answer. Biological knowledge about components of the system, its structure, interactions, and any available experimental results must be gathered. Different types of experimental data are analyzed and integrated. This information is then used to construct a mathematical model. Since different models emphasize different features, the choice of mathematical model depends on the questions being asked. Its structure will also depend on the system description: organismal, cellular, or molecular. Some systems will have unknown biological parameters and will require detailed information about kinetic constants or time course data in order to estimate model parameters. Various computational techniques are used to assess if the model is in accordance with experimental results, and if not, hypotheses underlying the model need to be refined, and different types of experiments might be proposed. This iterative process is repeated and the model is refined until it is accurately describes the relevant aspects of the system.
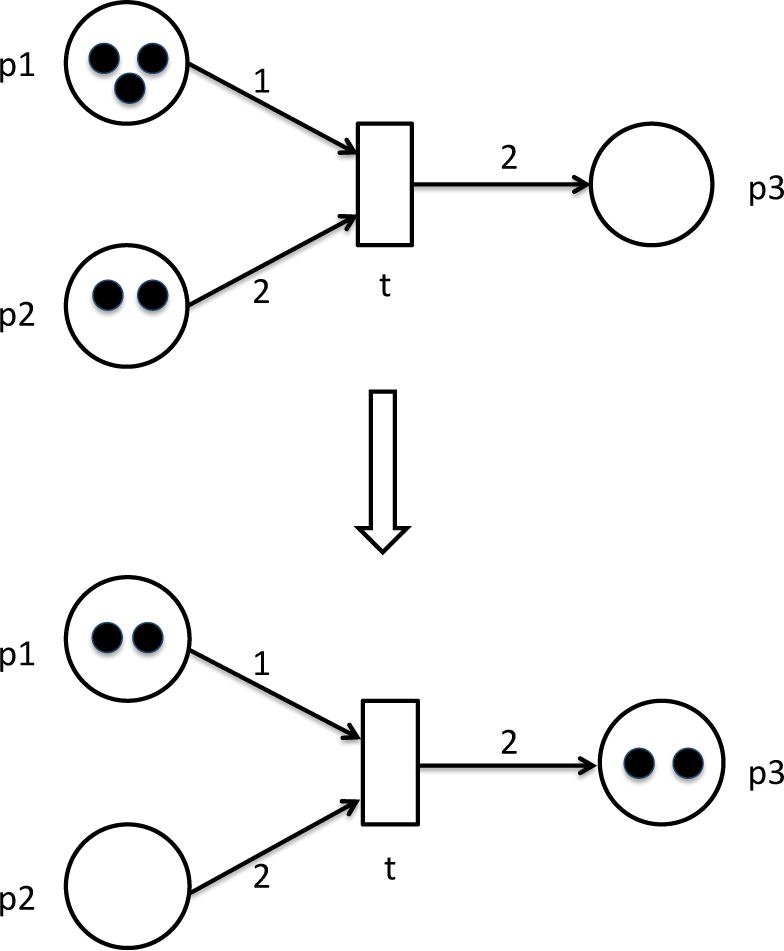
Initially input places p1 and p2 contain three and two tokens, respectively. By firing transition t one token will be removed from p1 and two tokens from p2. Transition t will consume tokens and place two of them into place p3. Transition t may fire since its preplaces p1 and p2 have sufficient number of tokens. After one firing step the marking of the net is changed: p1 has only two tokens left, p2 has no tokens, and p3 has two tokens. Transition t cannot fire anymore.
References
-
- Josephs HW. Absorption of iron as a problem in human physiology; a critical review. Blood. 1958;13(1):1–54. - PubMed
-
- Laufberger V. Sur la cristallisation de la ferritine. Soc Chim Biol. 1937;19:1575–1582.
-
- Feder JN, et al. A novel MHC class I-like gene is mutated in patients with hereditary haemochromatosis. Nat Genet. 1996;13(4):399–408. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
