

Chd1 is essential for the high transcriptional output and rapid growth of the mouse epiblast
- PMID: 25480920
- PMCID: PMC4299150
- DOI: 10.1242/dev.114843
Chd1 is essential for the high transcriptional output and rapid growth of the mouse epiblast
Abstract
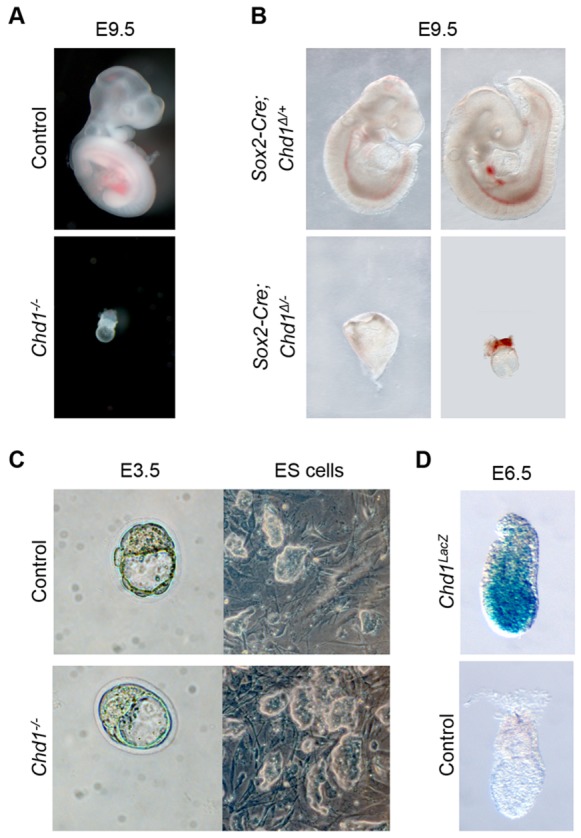
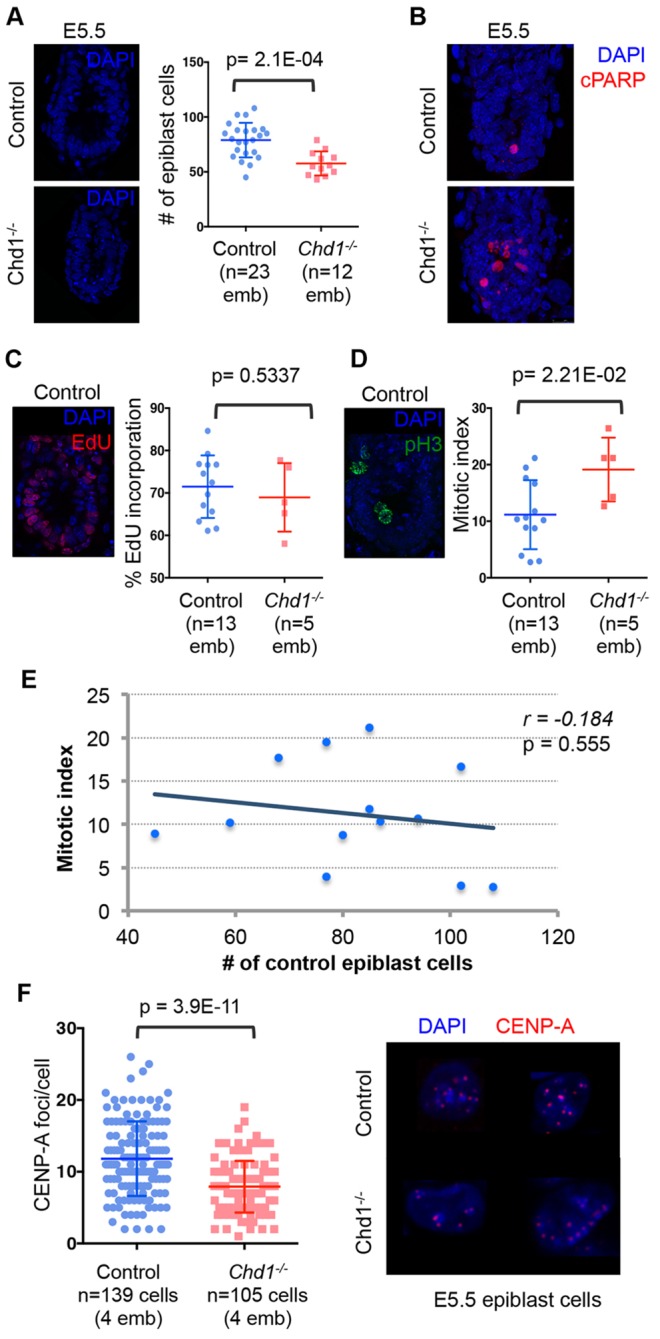
The pluripotent mammalian epiblast undergoes unusually fast cell proliferation. This rapid growth is expected to generate a high transcriptional demand, but the underlying mechanisms remain unknown. We show here that the chromatin remodeler Chd1 is required for transcriptional output and development of the mouse epiblast. Chd1(-/-) embryos exhibit proliferation defects and increased apoptosis, are smaller than controls by E5.5 and fail to grow, to become patterned or to gastrulate. Removal of p53 allows progression of Chd1(-/-) mutants only to E7.0-8.0, highlighting the crucial requirement for Chd1 during early post-implantation development. Chd1(-/-) embryonic stem cells (ESCs) have a self-renewal defect and a genome-wide reduction in transcriptional output at both known mRNAs and intergenic transcripts. These transcriptional defects were only uncovered when cell number-normalized approaches were used, and correlate with a lower engagement of RNAP II with transcribed genes in Chd1(-/-) ESCs. We further show that Chd1 directly binds to ribosomal DNA, and that both Chd1(-/-) epiblast cells in vivo and ESCs in vitro express significantly lower levels of ribosomal RNA. In agreement with these findings, mutant cells in vivo and in vitro exhibit smaller and more elongated nucleoli. Thus, the RNA output by both Pol I and II is reduced in Chd1(-/-) cells. Our data indicate that Chd1 promotes a globally elevated transcriptional output required to sustain the distinctly rapid growth of the mouse epiblast.
Keywords: Chromatin; Epiblast proliferation; Epigenetics; Pluripotency; Transcription.
© 2015. Published by The Company of Biologists Ltd.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
