

Targeted mutagenesis of aryl hydrocarbon receptor 2a and 2b genes in Atlantic killifish (Fundulus heteroclitus)
- PMID: 25481785
- PMCID: PMC4272816
- DOI: 10.1016/j.aquatox.2014.11.016
Targeted mutagenesis of aryl hydrocarbon receptor 2a and 2b genes in Atlantic killifish (Fundulus heteroclitus)
Abstract
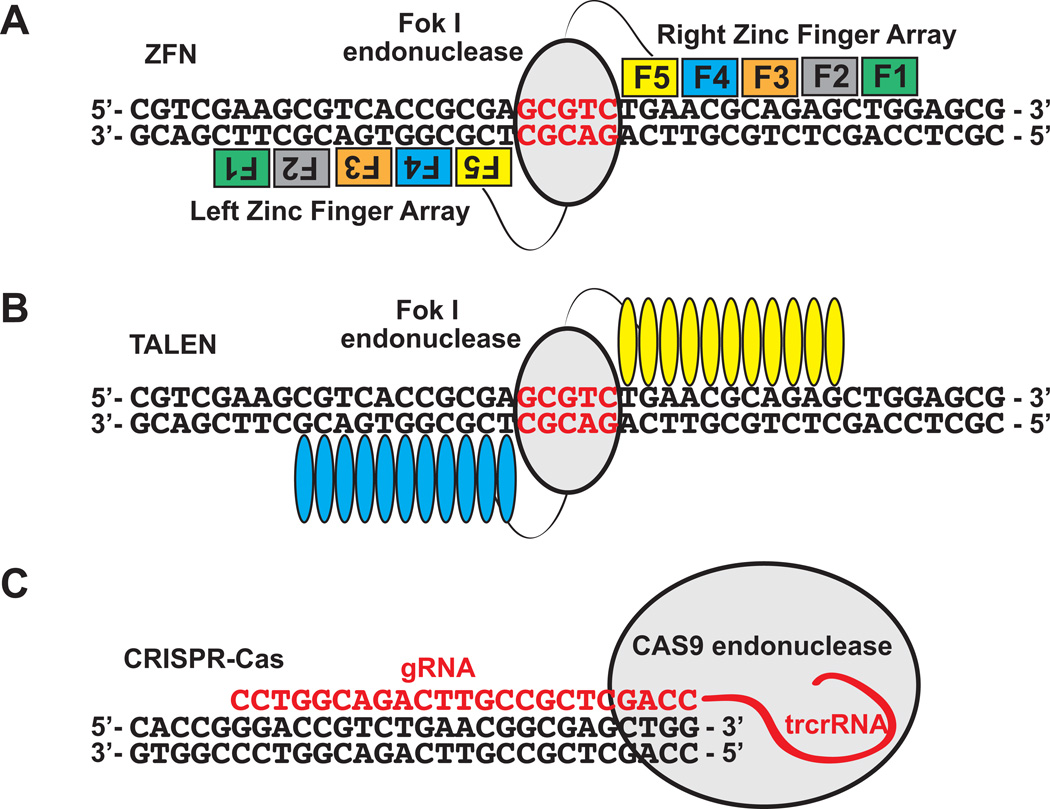
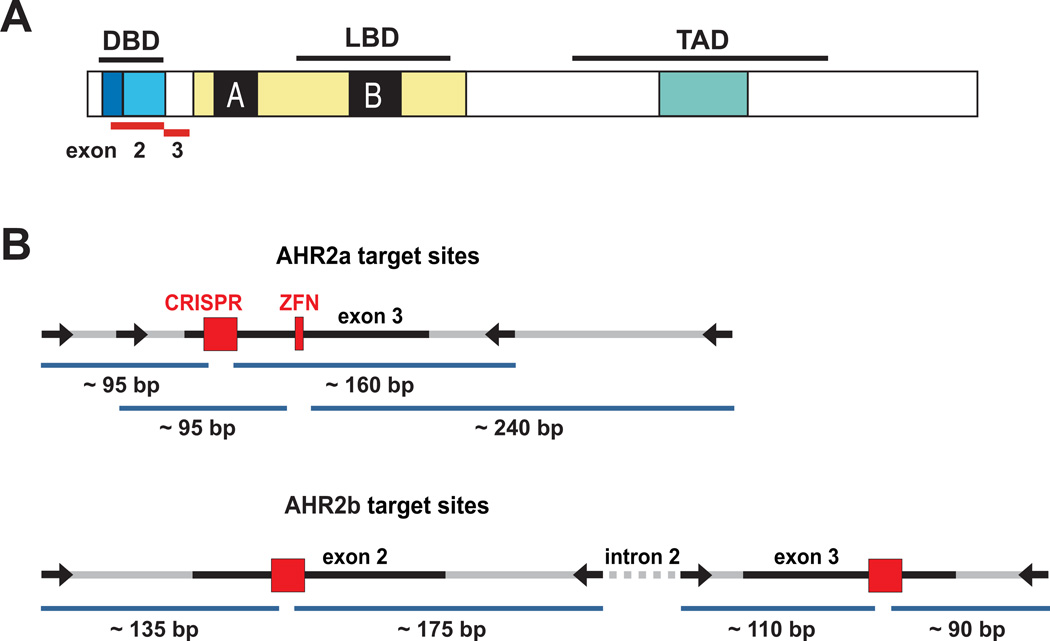
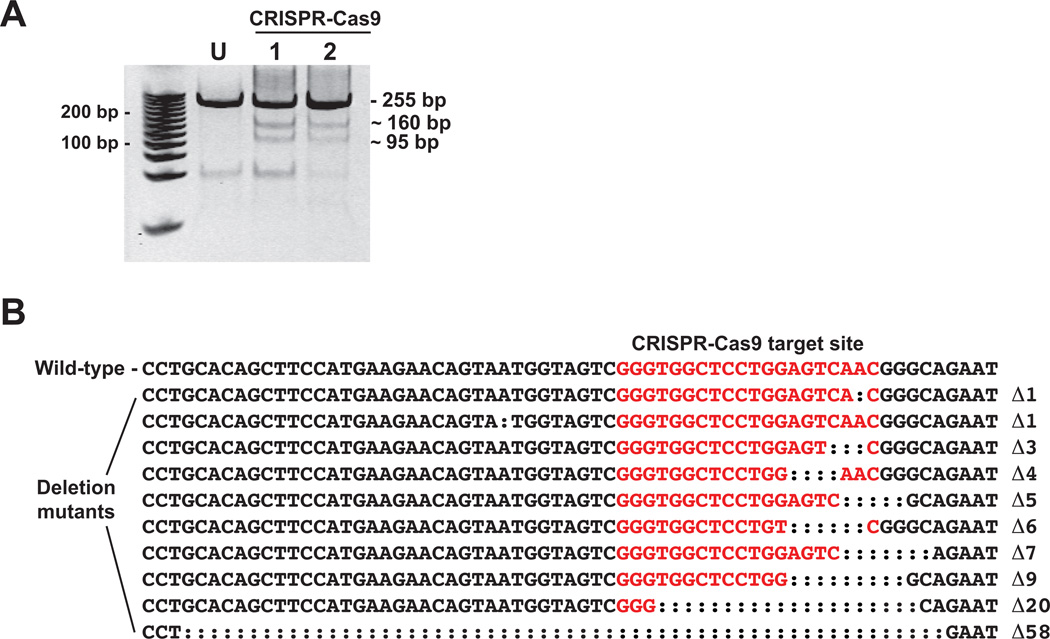
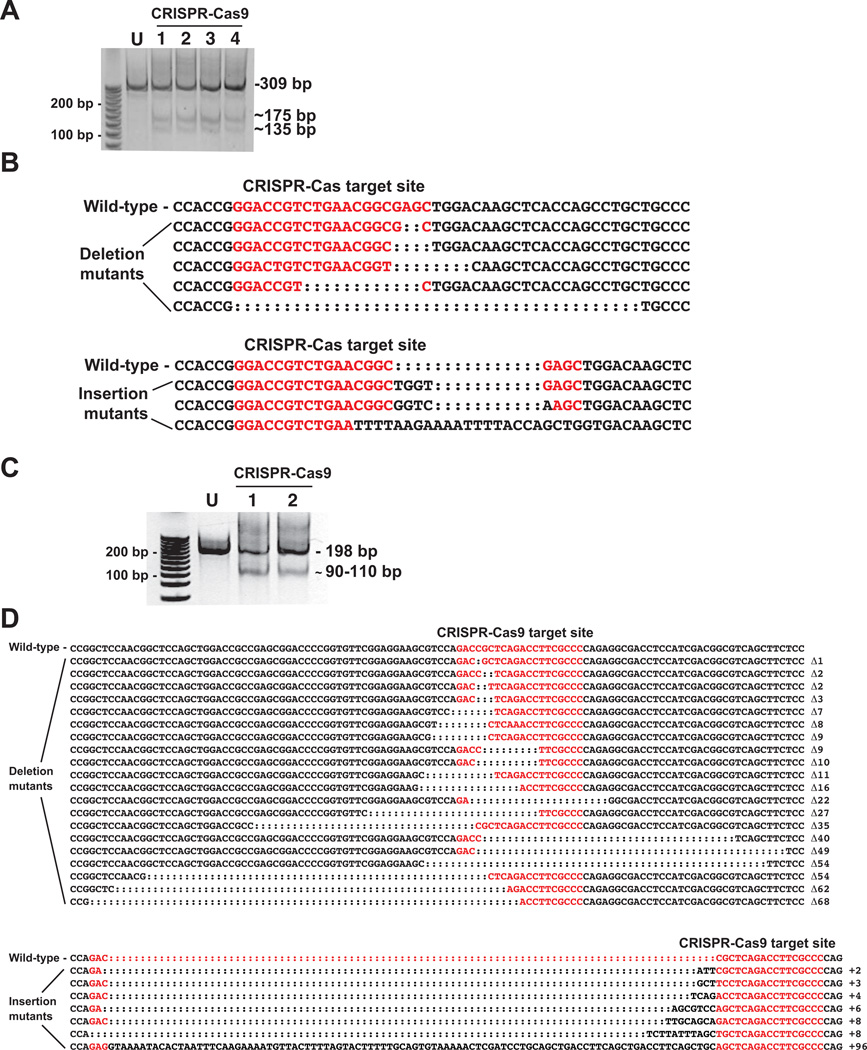
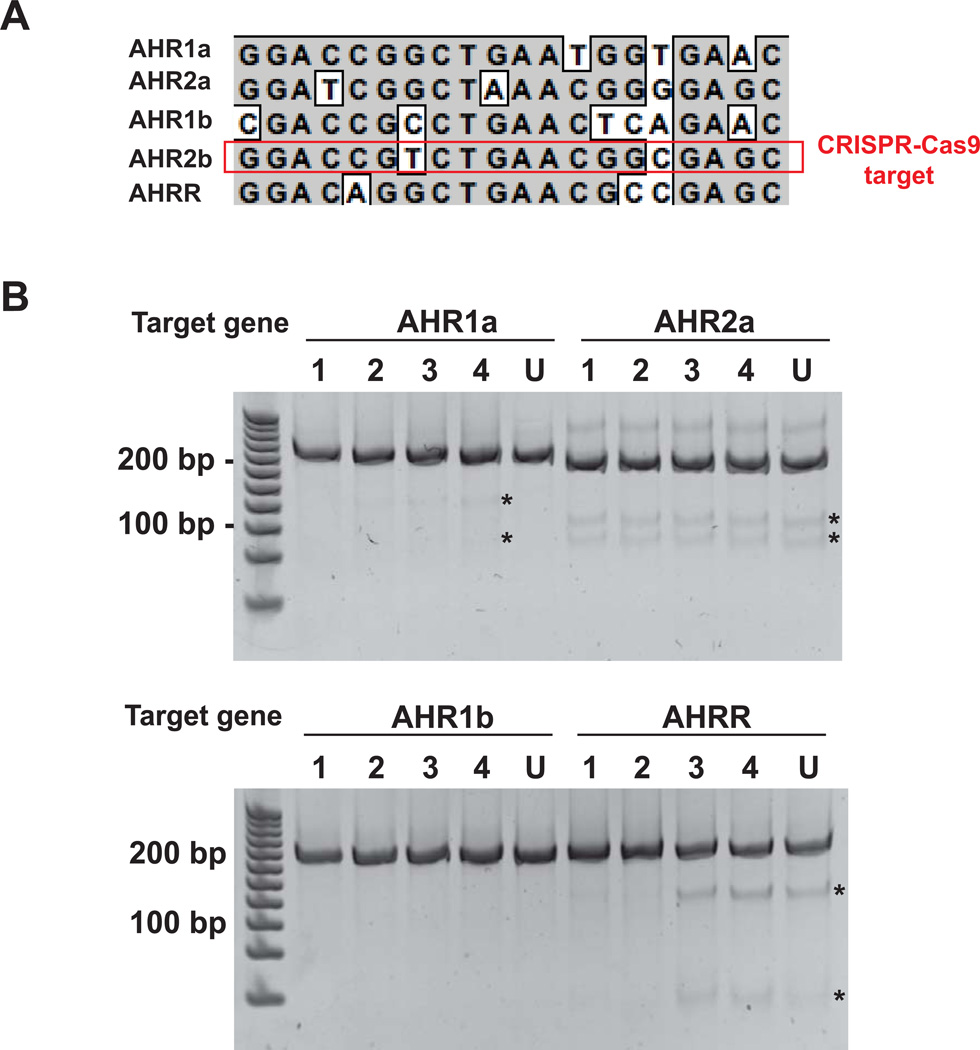
Understanding molecular mechanisms of toxicity is facilitated by experimental manipulations, such as disruption of function by gene targeting, that are especially challenging in non-standard model species with limited genomic resources. While loss-of-function approaches have included gene knock-down using morpholino-modified oligonucleotides and random mutagenesis using mutagens or retroviruses, more recent approaches include targeted mutagenesis using zinc finger nuclease (ZFN), transcription activator-like effector nuclease (TALENs) and clustered regularly interspaced short palindromic repeats (CRISPR)-Cas9 technology. These latter methods provide more accessible opportunities to explore gene function in non-traditional model species. To facilitate evaluation of toxic mechanisms for important categories of aryl hydrocarbon pollutants, whose actions are known to be receptor mediated, we used ZFN and CRISPR-Cas9 approaches to generate aryl hydrocarbon receptor 2a (AHR2a) and AHR2b gene mutations in Atlantic killifish (Fundulus heteroclitus) embryos. This killifish is a particularly valuable non-traditional model, with multiple paralogs of AHR whose functions are not well characterized. In addition, some populations of this species have evolved resistance to toxicants such as halogenated aromatic hydrocarbons. AHR-null killifish will be valuable for characterizing the role of the individual AHR paralogs in evolved resistance, as well as in normal development. We first used five-finger ZFNs targeting exons 1 and 3 of AHR2a. Subsequently, CRISPR-Cas9 guide RNAs were designed to target regions in exon 2 and 3 of AHR2a and AHR2b. We successfully induced frameshift mutations in AHR2a exon 3 with ZFN and CRISPR-Cas9 guide RNAs, with mutation frequencies of 10% and 16%, respectively. In AHR2b, mutations were induced using CRISPR-Cas9 guide RNAs targeting sites in both exon 2 (17%) and exon 3 (63%). We screened AHR2b exon 2 CRISPR-Cas9-injected embryos for off-target effects in AHR paralogs. No mutations were observed in closely related AHR genes (AHR1a, AHR1b, AHR2a, AHRR) in the CRISPR-Cas9-injected embryos. Overall, our results demonstrate that targeted genome-editing methods are efficient in inducing mutations at specific loci in embryos of a non-traditional model species, without detectable off-target effects in paralogous genes.
Keywords: Adaptation; CRISPR-Cas9; Gene knock-outs; Mummichog; Non-model organisms; Zinc finger nucleases.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures

References
-
- Aluru N, Karchner SI, Jenny MJ, Franks D, Hahn ME. Understanding the physiological role of aryl hydrocarbon receptor repressor (AHRR) using gene knock-down and targeted mutagenesis in zebrafish. Annual Meeting of Society of Toxicology; San Antonio, Texas. 2013.
-
- Ansai S, Inohaya K, Yoshiura Y, Schartl M, Uemura N, Takahashi R, Kinoshita M. Design, evaluation, and screening methods for efficient targeted mutagenesis with transcription activator-like effector nucleases in medaka. Dev Growth Differ. 2014;56:98–107. - PubMed
-
- Ansai S, Ochiai H, Kanie Y, Kamei Y, Gou Y, Kitano T, Yamamoto T, Kinoshita M. Targeted disruption of exogenous EGFP gene in medaka using zinc-finger nucleases. Dev Growth Differ. 2012;54:546–556. - PubMed
-
- Armstrong PB, Child JS. Stages in Normal Development of Fundulus heteroclitus. Biol Bull. 1965;128:143–168.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
