

Mesenchymal chemotaxis requires selective inactivation of myosin II at the leading edge via a noncanonical PLCγ/PKCα pathway
- PMID: 25482883
- PMCID: PMC4276478
- DOI: 10.1016/j.devcel.2014.10.024
Mesenchymal chemotaxis requires selective inactivation of myosin II at the leading edge via a noncanonical PLCγ/PKCα pathway
Abstract
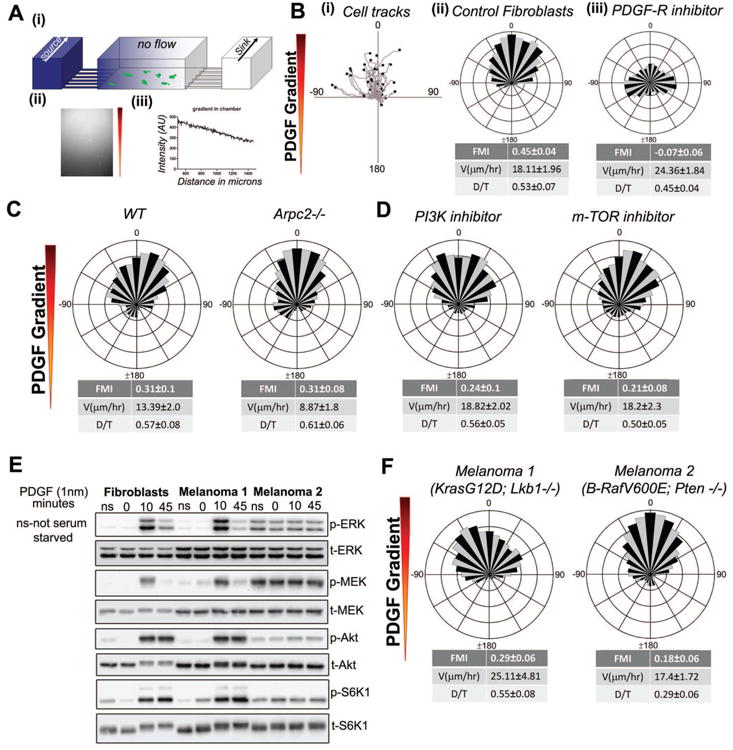
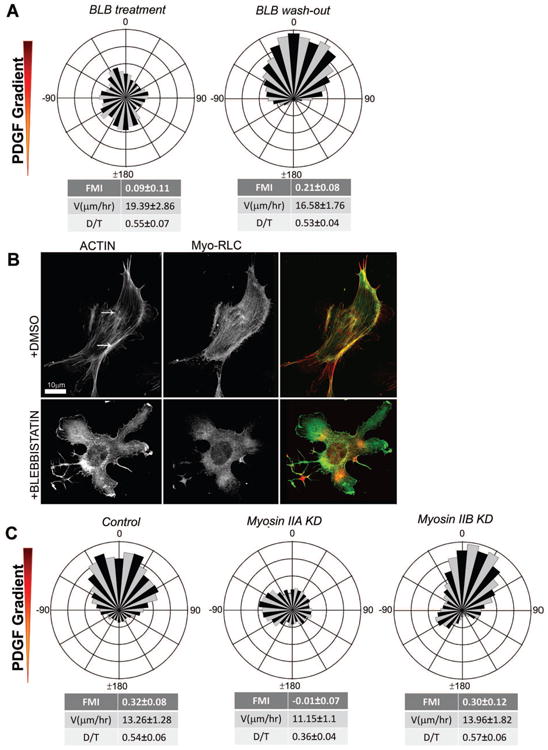
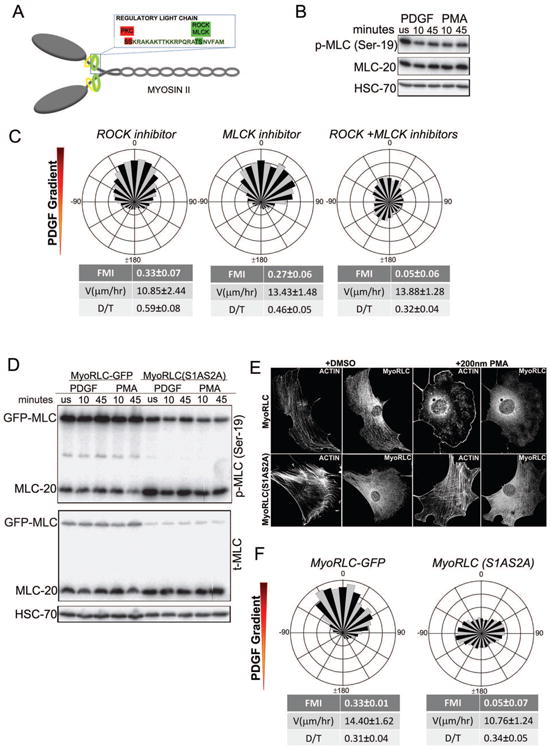
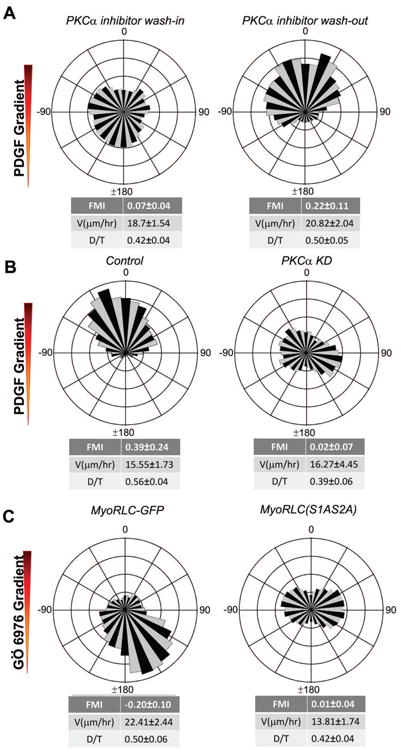
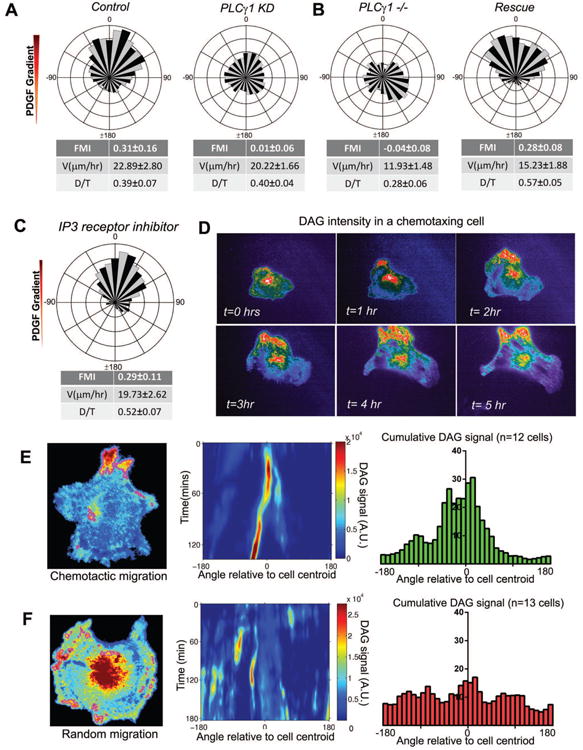
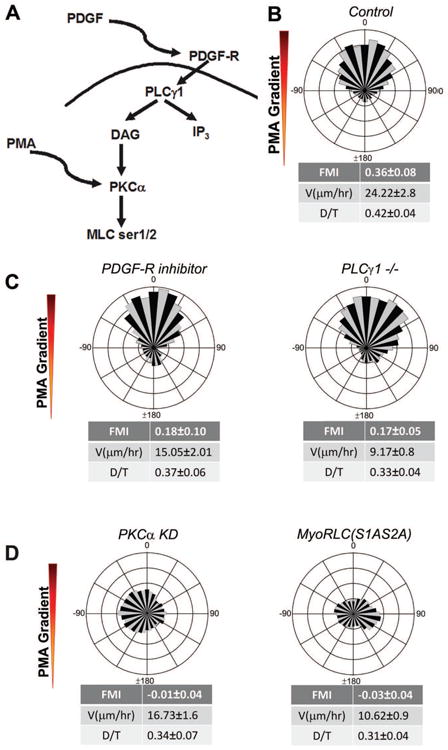
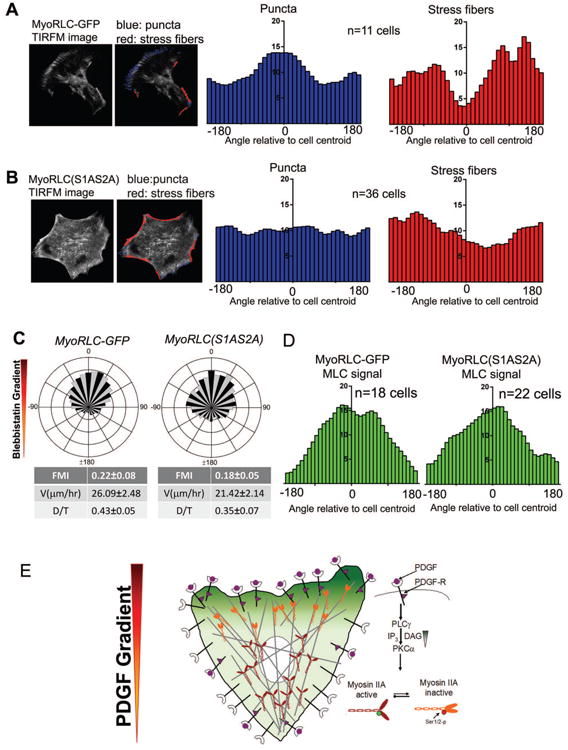
Chemotaxis, migration toward soluble chemical cues, is critical for processes such as wound healing and immune surveillance and is exhibited by various cell types, from rapidly migrating leukocytes to slow-moving mesenchymal cells. To study mesenchymal chemotaxis, we observed cell migration in microfluidic chambers that generate stable gradients of platelet-derived growth factor (PDGF). Surprisingly, we found that pathways implicated in amoeboid chemotaxis, such as PI3K and mammalian target of rapamycin signaling, are dispensable for PDGF chemotaxis. Instead, we find that local inactivation of Myosin IIA, through a noncanonical Ser1/2 phosphorylation of the regulatory light chain, is essential. This site is phosphorylated by PKCα, which is activated by an intracellular gradient of diacylglycerol generated by PLCγ. Using a combination of live imaging and gradients of activators/inhibitors in the microfluidic chambers, we demonstrate that this signaling pathway and subsequent inhibition of Myosin II activity at the leading edge are required for mesenchymal chemotaxis.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Unleashing mesenchymal chemotaxis.Dev Cell. 2014 Dec 22;31(6):669-70. doi: 10.1016/j.devcel.2014.12.005. Dev Cell. 2014. PMID: 25535912
References
-
- Bear J, Loureiro J, Libova I, Fassler R, Wehland J, Gertler F. Negative regulation of fibroblast motility by Ena/VASP proteins. Cell. 2000;101:717–728. - PubMed
-
- Bear JE, Svitkina TM, Krause M, Schafer DA, Loureiro JJ, Strasser GA, Maly IV, Chaga OY, Cooper JA, Borisy GG, et al. Antagonism between Ena/VASP Proteins and Actin Filament Capping Regulates Fibroblast Motility. Cell. 2002;109:509–521. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
