

Unresolved issues in theories of autoimmune disease using myocarditis as a framework
- PMID: 25484004
- PMCID: PMC4417457
- DOI: 10.1016/j.jtbi.2014.11.022
Unresolved issues in theories of autoimmune disease using myocarditis as a framework
Abstract
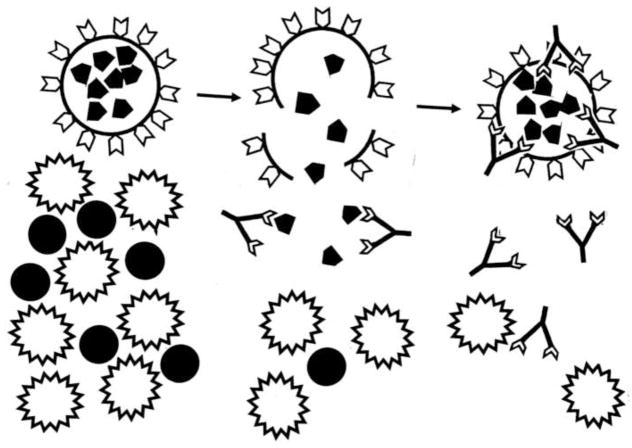
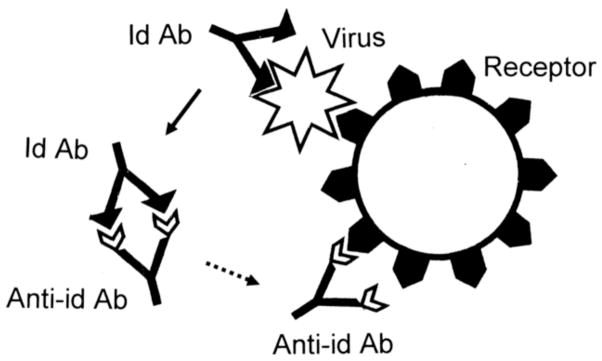
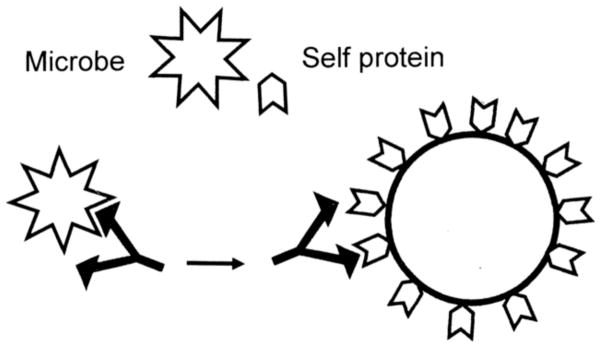
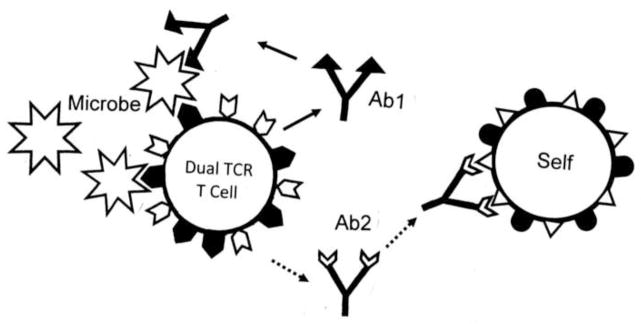
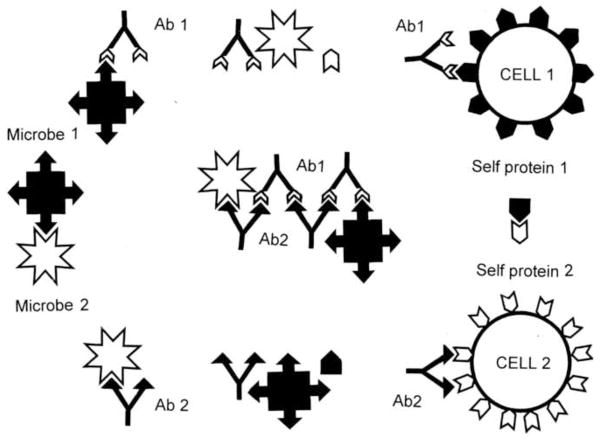
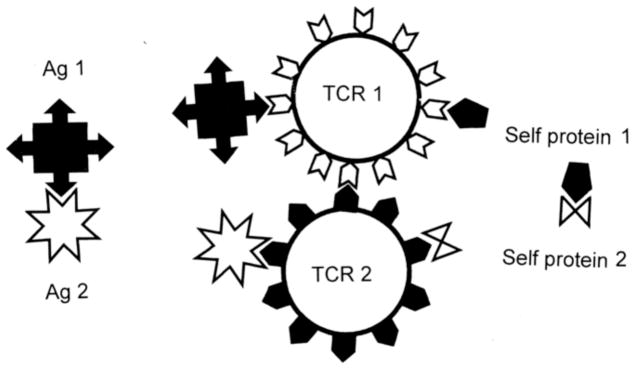
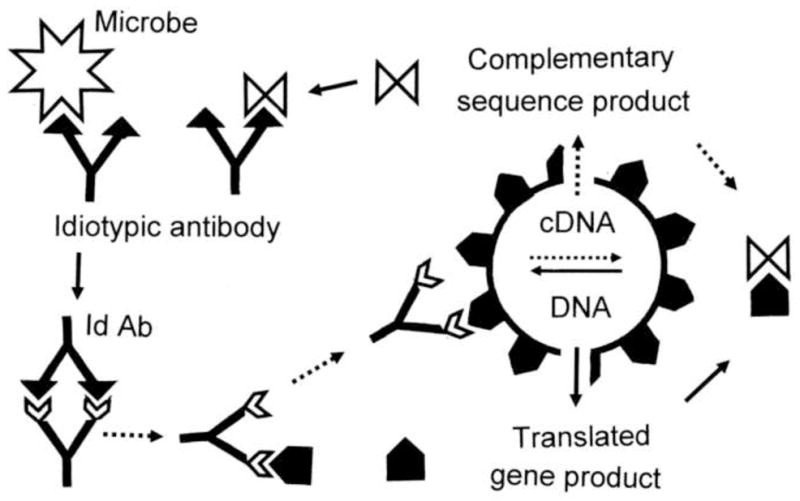
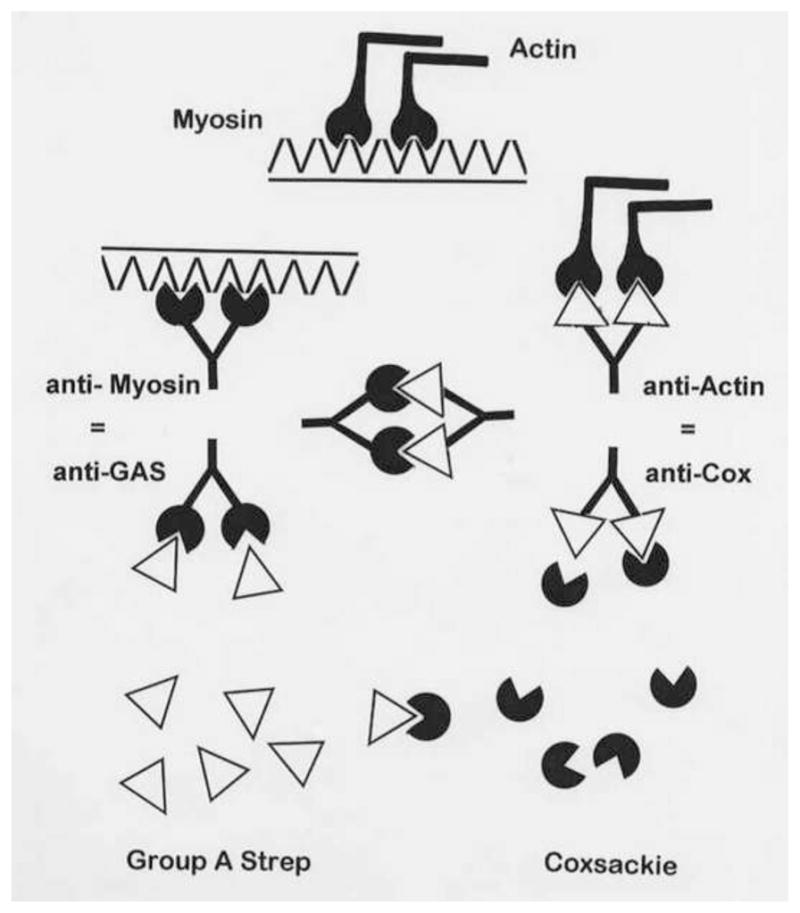
Many theories of autoimmune disease have been proposed since the discovery that the immune system can attack the body. These theories include the hidden or cryptic antigen theory, modified antigen theory, T cell bypass, T cell-B cell mismatch, epitope spread or drift, the bystander effect, molecular mimicry, anti-idiotype theory, antigenic complementarity, and dual-affinity T cell receptors. We critically review these theories and relevant mathematical models as they apply to autoimmune myocarditis. All theories share the common assumption that autoimmune diseases are triggered by environmental factors such as infections or chemical exposure. Most, but not all, theories and mathematical models are unifactorial assuming single-agent causation of disease. Experimental and clinical evidence and mathematical models exist to support some aspects of most theories, but evidence/models that support one theory almost invariably supports other theories as well. More importantly, every theory (and every model) lacks the ability to account for some key autoimmune disease phenomena such as the fundamental roles of innate immunity, sex differences in disease susceptibility, the necessity for adjuvants in experimental animal models, and the often paradoxical effect of exposure timing and dose on disease induction. We argue that a more comprehensive and integrated theory of autoimmunity associated with new mathematical models is needed and suggest specific experimental and clinical tests for each major theory that might help to clarify how they relate to clinical disease and reveal how theories are related.
Keywords: Antigenic complementarity; Autoimmune disease theories and modeling; Infection; Myocarditis; Sex differences.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
