

Autophagy facilitates secretion and protects against degeneration of the Harderian gland
- PMID: 25484081
- PMCID: PMC4502725
- DOI: 10.4161/15548627.2014.978221
Autophagy facilitates secretion and protects against degeneration of the Harderian gland
Abstract
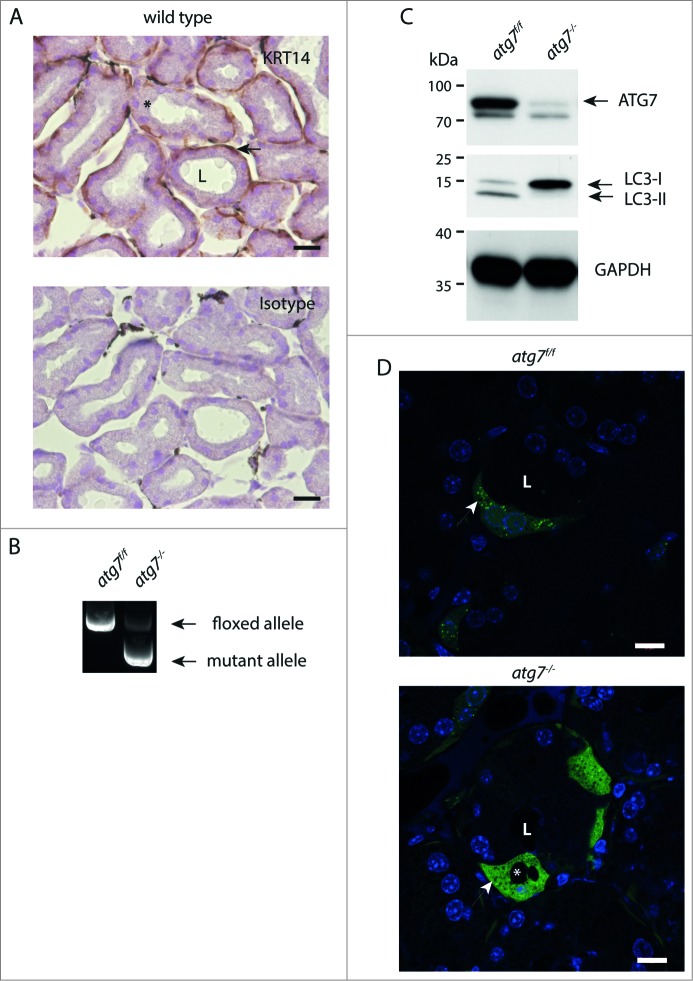
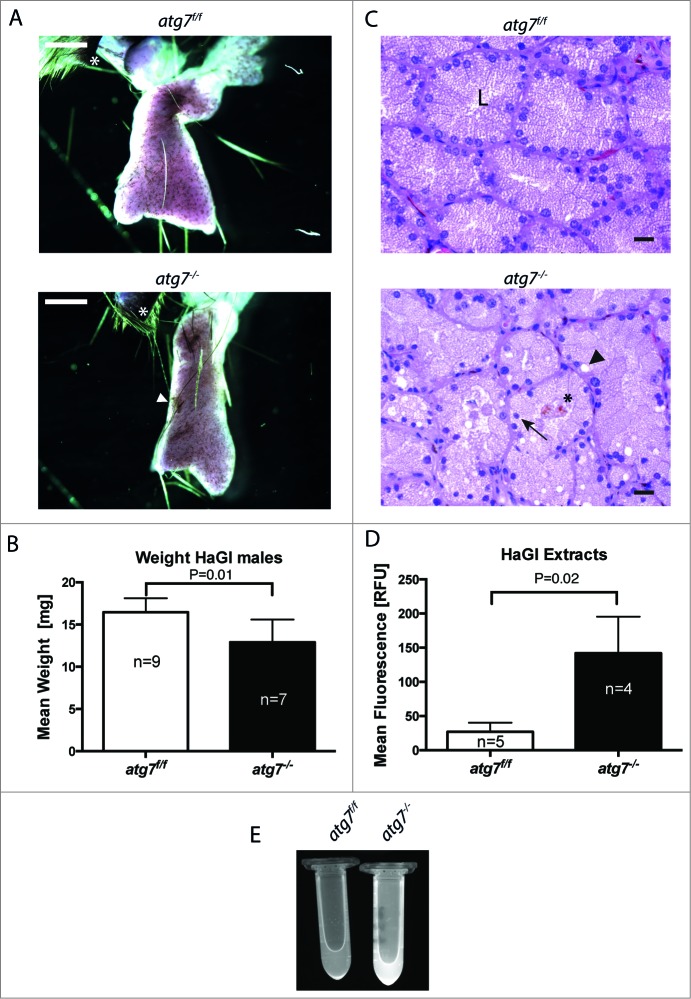
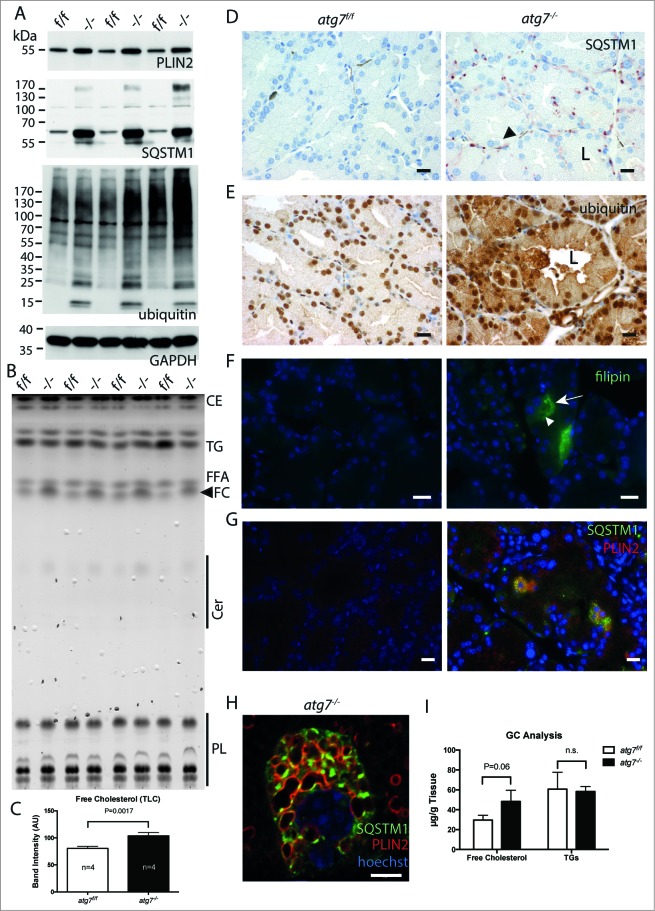
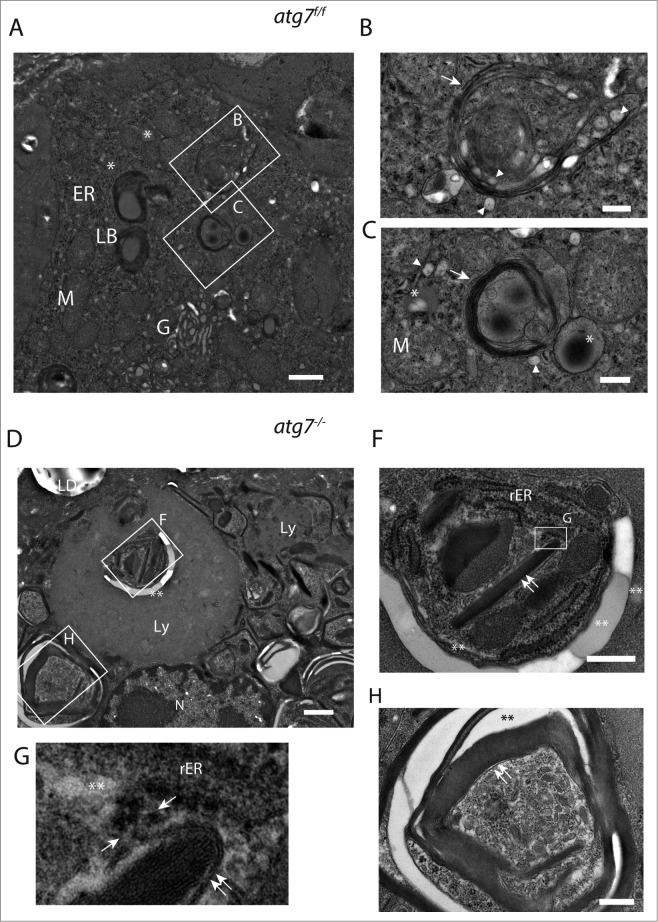
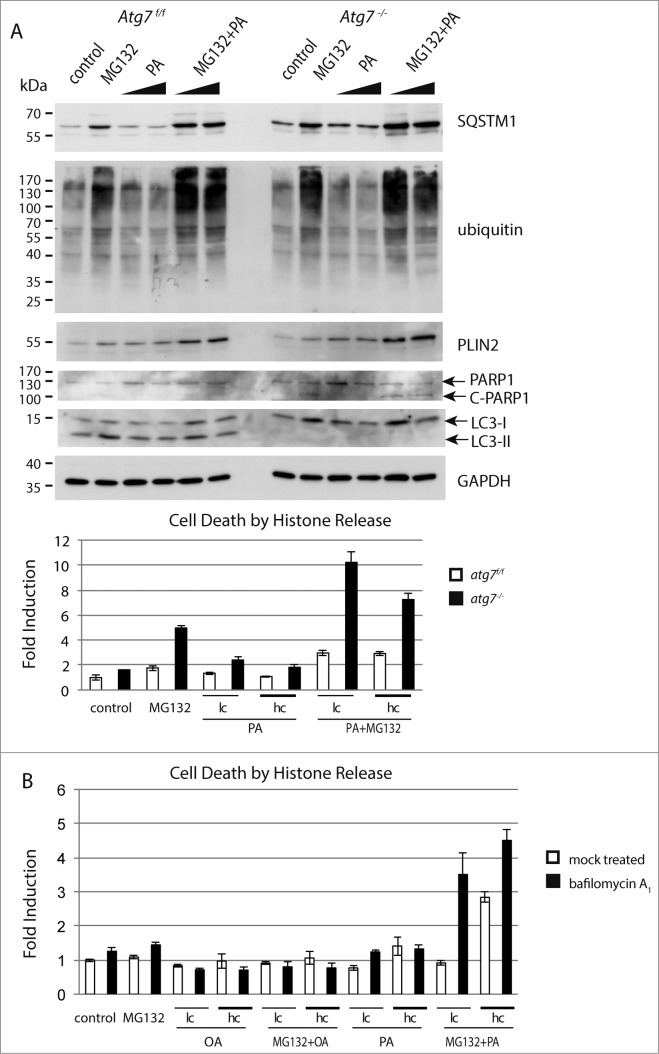
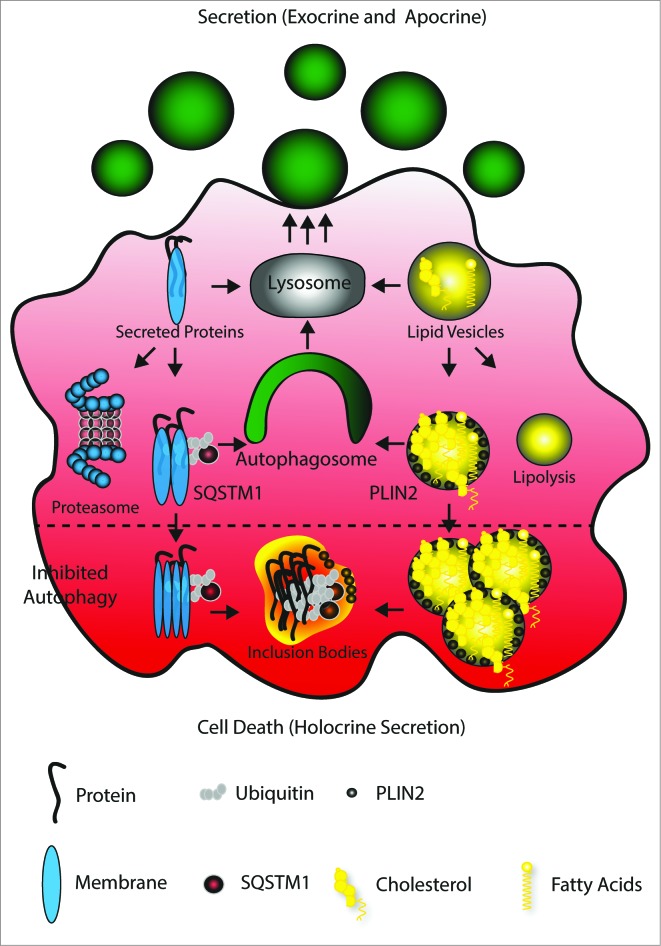
The epithelial derived Harderian gland consists of 2 types of secretory cells. The more numerous type A cells are responsible for the secretion of lipid droplets, while type B cells produce dark granules of multilamellar bodies. The process of autophagy is constitutively active in the Harderian gland, as confirmed by our analysis of LC3 processing in GFP-LC3 transgenic mice. This process is compromised by epithelial deletion of Atg7. Morphologically, the Atg7 mutant glands are hypotrophic and degenerated, with highly vacuolated cells and pyknotic nuclei. The mutant glands accumulate lipid droplets coated with PLIN2 (perilipin 2) and contain deposits of cholesterol, ubiquitinated proteins, SQSTM1/p62 (sequestosome 1) positive aggregates and other metabolic products such as porphyrin. Immunofluorescence stainings show that distinct cells strongly aggregate both proteins and lipids. Electron microscopy of the Harderian glands reveals that its organized structure is compromised, and the presence of large intracellular lipid droplets and heterologous aggregates. We attribute the occurrence of large vacuoles to a malfunction in the formation of multilamellar bodies found in the less abundant type B Harderian gland cells. This defect causes the formation of large tertiary lysosomes of heterologous content and is accompanied by the generation of tight lamellar stacks of endoplasmic reticulum in a pseudo-crystalline form. To test the hypothesis that lipid and protein accumulation is the cause for the degeneration in autophagy-deficient Harderian glands, epithelial cells were treated with a combination of the proteasome inhibitor and free fatty acids, to induce aggregation of misfolded proteins and lipid accumulation, respectively. The results show that lipid accumulation indeed enhanced the toxicity of misfolded proteins and that this was even more pronounced in autophagy-deficient cells. Thus, we conclude autophagy controls protein and lipid catabolism and anabolism to facilitate bulk production of secretory vesicles of the Harderian gland.
Keywords: Atg12, autophagy related 12; Atg7, autophagy related 7; BCA, bicinchoninic acid assay; BODIPY, boron-dipyrromethene fluorescent dye; BSA, bovine serum albumin; Cre, Cre recombinase; DMSO, dimethyl sulfoxide; ELISA, enzyme-linked immunosorbent assay; ER, edoplasmic reticulum; FC, free cholesterol; GFP, green fluorescent protein; HaGl, Harderian gland; Harderian gland; KLICK, keratosis lineariz with ichthyosis congenita and sclerosing keratoderma; KRT14, Keratin 14; LD, Lipid drops; LSM, laser scanning microscope; MAP1LC3A/B (LC3), microtubule-associated protein 1 light chain 3 α/β; MG132; MG312, synthetic peptide Z-Leu-Leu-Leu-al; ORO, oil red O; PARP, poly (ADP-ribose) polymerase; PCR, polymerase chain reaction; PLIN2, perilipin 2; RFU, relative fluorecent units; SQSTM1, sequestosome 1/p62; SQSTM1/p62; TBS-T, Tris buffered saline with Tween 20; TLC, thin layer chromatography; UV, ultraviolet; aggregates; aggresome; autophagy; cholesterol; degenerative diseases; f, floxed; keratinocytes; lipotoxicity; lysosome; multilamellar bodies; palmitate; perilipin 2/adipophilin; proteasome inhibitor.
Figures
References
-
- Brookheart RT, Michel CI, Schaffer JE. As a matter of fat. Cell Metab 2009; 10:9-12; PMID:19583949; http://dx.doi.org/10.1016/j.cmet.2009.03.011 - DOI - PMC - PubMed
-
- Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell 2012; 149:274-93; PMID:22500797; http://dx.doi.org/10.1016/j.cell.2012.03.017 - DOI - PMC - PubMed
-
- Ouellet J, Barral Y. Organelle segregation during mitosis: lessons from asymmetrically dividing cells. J Cell Biol 2012; 196:305-13; PMID:22312002; http://dx.doi.org/10.1083/jcb.201102078 - DOI - PMC - PubMed
-
- Lin H. Cell biology of stem cells: an enigma of asymmetry and self-renewal. J Cell Biol 2008; 180:257-60; PMID:18227277; http://dx.doi.org/10.1083/jcb.200712159 - DOI - PMC - PubMed
-
- Lippens S, Denecker G, Ovaere P, Vandenabeele P, Declercq W. Death penalty for keratinocytes: apoptosis versus cornification. Cell Death Differ 2005; 12 Suppl 2:1497-508; PMID:16247497; http://dx.doi.org/10.1038/sj.cdd.4401722 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous