

Critical determinants of mitochondria-associated neutral sphingomyelinase (MA-nSMase) for mitochondrial localization
- PMID: 25484313
- PMCID: PMC4435939
- DOI: 10.1016/j.bbagen.2014.11.019
Critical determinants of mitochondria-associated neutral sphingomyelinase (MA-nSMase) for mitochondrial localization
Abstract
Background: A novel murine mitochondria-associated neutral sphingomyelinase (MA-nSMase) has been recently cloned and partially characterized. The subcellular localization of the enzyme was found to be predominant in mitochondria. In this work, the determinants of mitochondrial localization and its topology were investigated.
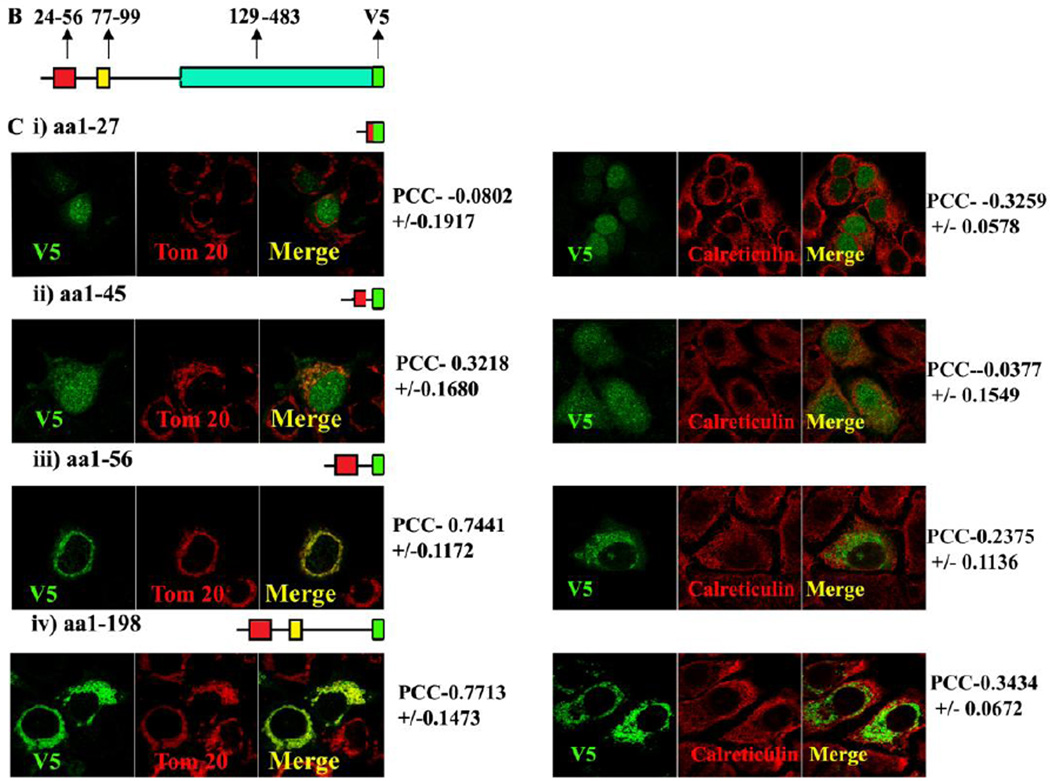
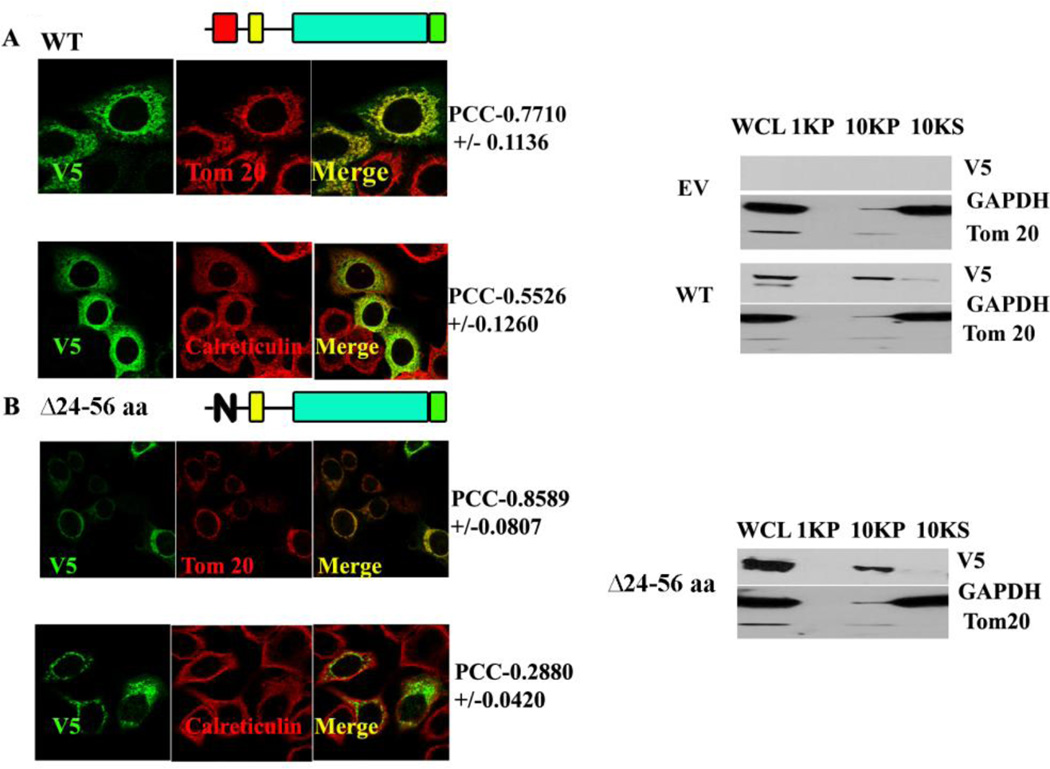
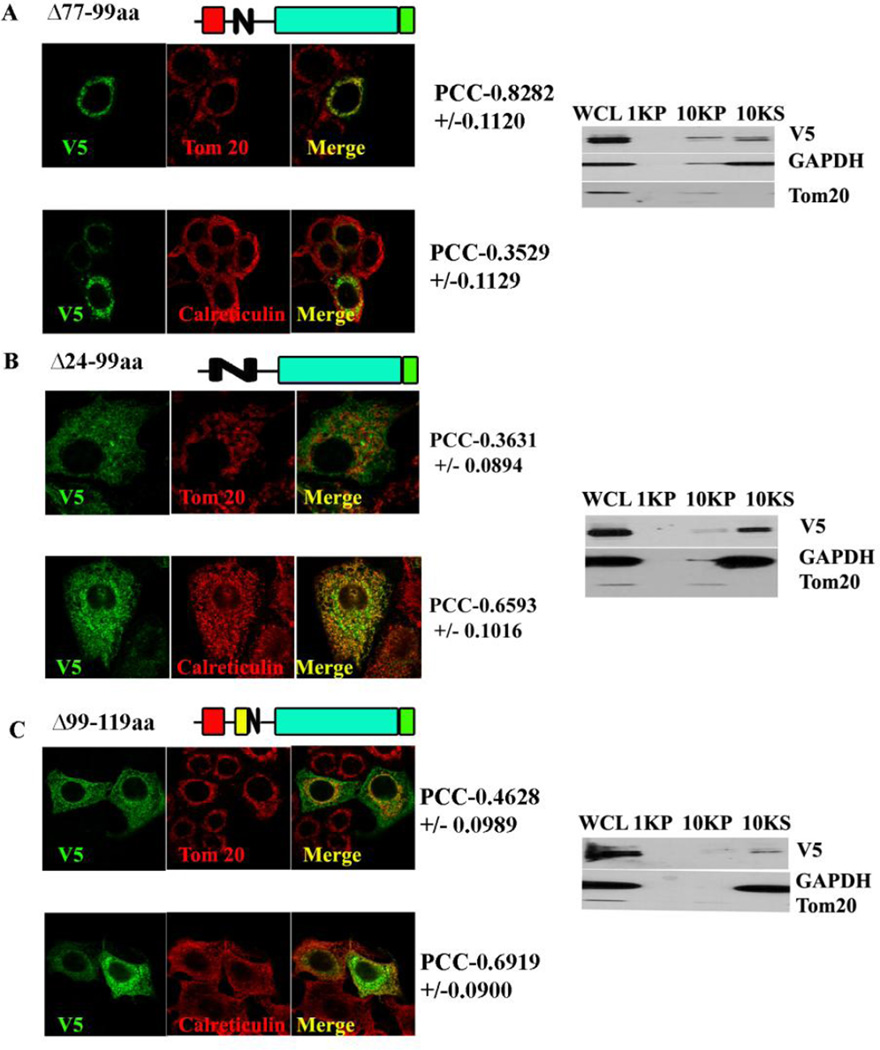
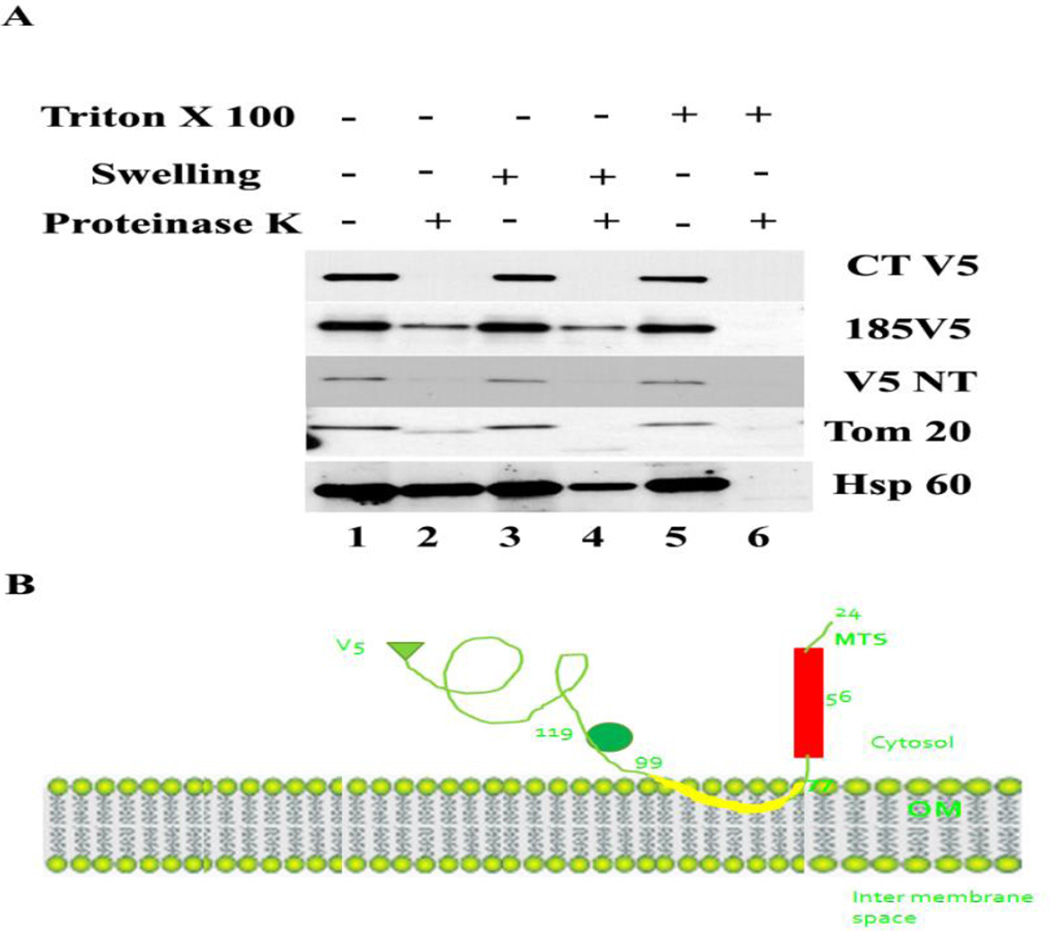
Methods: MA-nSMase mutants lacking consecutive regions and fusion proteins of GFP with truncated MA-nSMase regions were constructed and expressed in MCF-7 cells. Its localization was analyzed using confocal microscopy and sub-cellular fractionation methods. The sub-mitochondrial localization of MA-nSMase was determined using protease protection assay on isolated mitochondria.
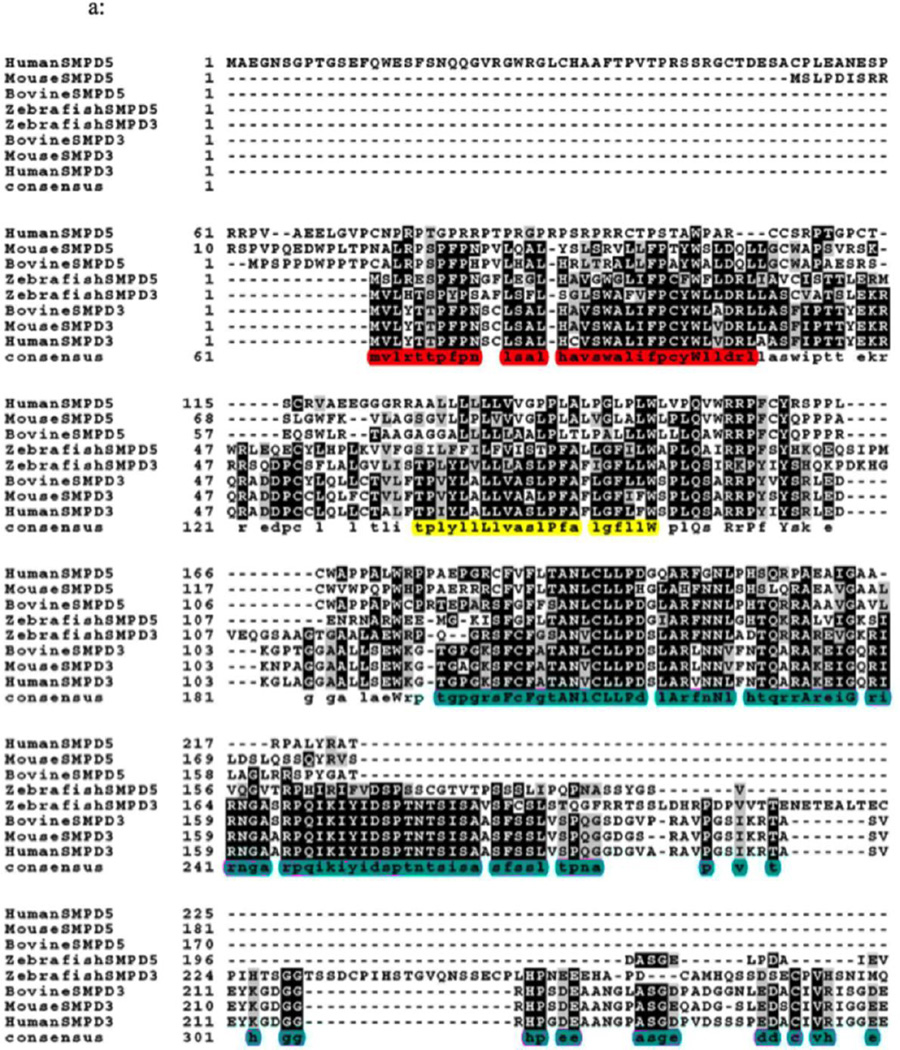
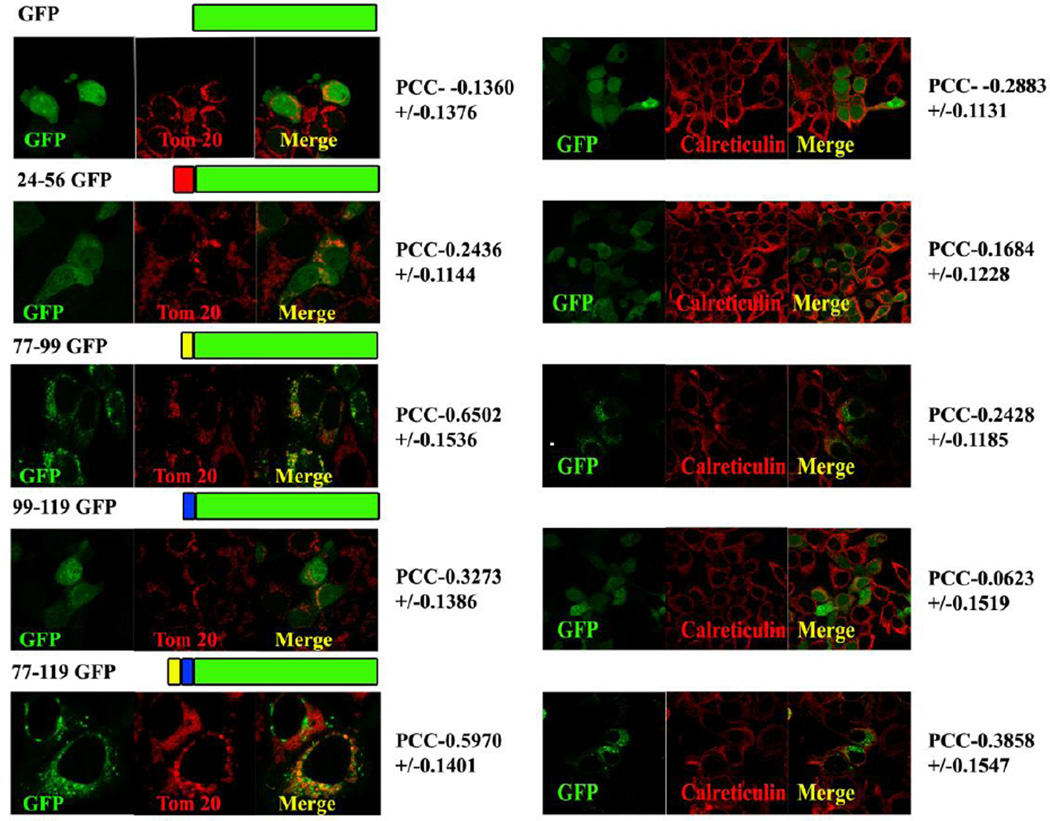
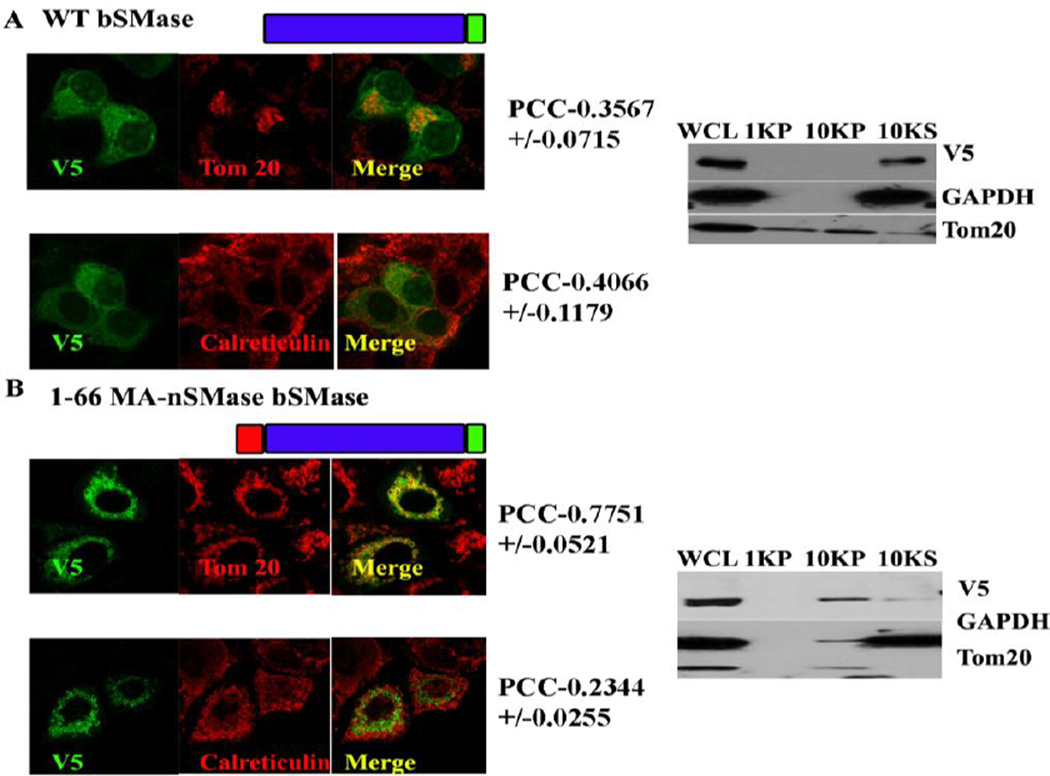
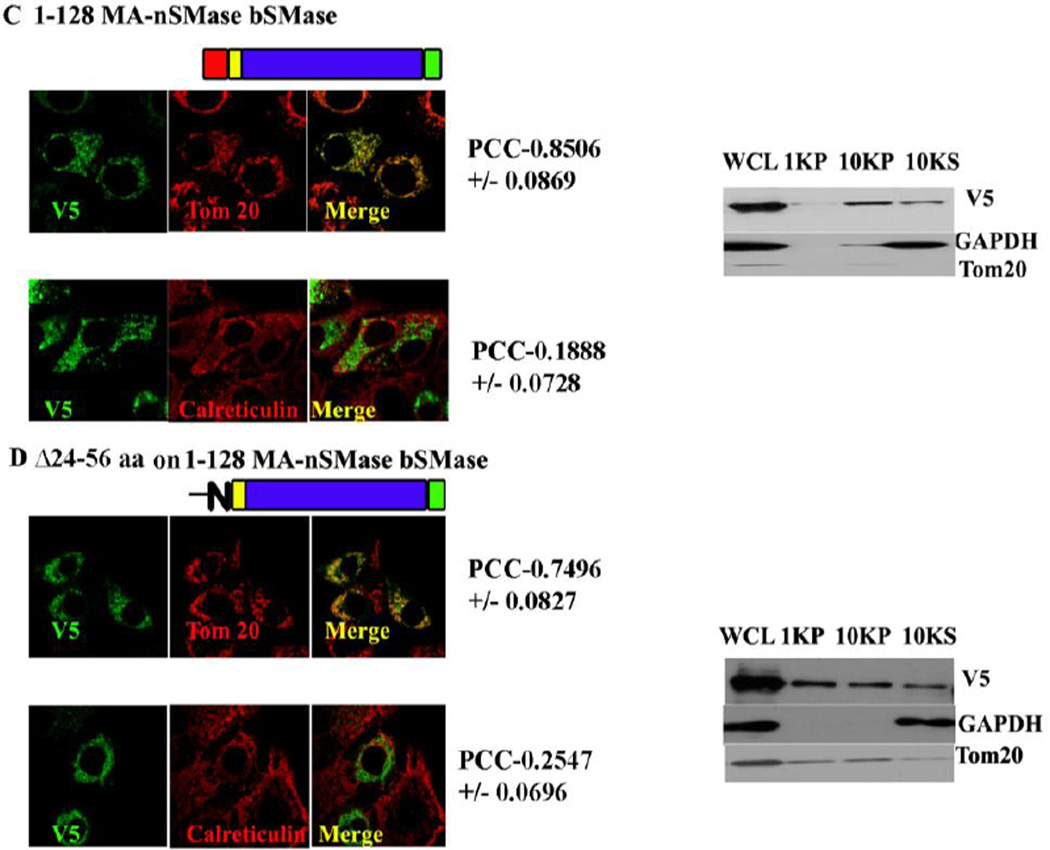
Results: The results initially showed that a putative mitochondrial localization signal (MLS), homologous to an MLS in the zebra-fish mitochondrial SMase is not necessary for the mitochondrial localization of the murine MA-nSMase. Evidence is provided to the presence of two regions in MA-nSMase that are sufficient for mitochondrial localization: a signal sequence (amino acids 24-56) that is responsible for the mitochondrial localization and an additional 'signal-anchor' sequence (amino acids 77-99) that anchors the protein to the mitochondrial membrane. This protein is topologically located in the outer mitochondrial membrane where both the C and N-termini remain exposed to the cytosol.
Conclusions: MA-nSMase is a membrane anchored protein with a MLS and a signal-anchor sequence at its N-terminal to localize it to the outer mitochondrial membrane.
General significance: Mitochondrial sphingolipids have been reported to play a critical role in cellular viability. This study opens a new window to investigate their cellular functions, and to define novel therapeutic targets.
Keywords: MA-nSMase; MLS; Mitochondrion; Sphingolipid; Sphingomyelinase.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Identification and characterization of murine mitochondria-associated neutral sphingomyelinase (MA-nSMase), the mammalian sphingomyelin phosphodiesterase 5.J Biol Chem. 2010 Jun 4;285(23):17993-8002. doi: 10.1074/jbc.M110.102988. Epub 2010 Apr 8. J Biol Chem. 2010. PMID: 20378533 Free PMC article.
-
A novel mitochondrial sphingomyelinase in zebrafish cells.J Biol Chem. 2009 Jul 24;284(30):20349-63. doi: 10.1074/jbc.M109.004580. Epub 2009 May 8. J Biol Chem. 2009. PMID: 19429680 Free PMC article.
-
Nuclear localization of neutral sphingomyelinase 1: biochemical and immunocytochemical analyses.J Cell Sci. 2001 Oct;114(Pt 20):3727-36. doi: 10.1242/jcs.114.20.3727. J Cell Sci. 2001. PMID: 11707524
-
[Structure and function of sphingomyelinase].Yakugaku Zasshi. 2009 Oct;129(10):1233-8. doi: 10.1248/yakushi.129.1233. Yakugaku Zasshi. 2009. PMID: 19797879 Review. Japanese.
-
Thematic review series: sphingolipids. ISC1 (inositol phosphosphingolipid-phospholipase C), the yeast homologue of neutral sphingomyelinases.J Lipid Res. 2008 May;49(5):922-8. doi: 10.1194/jlr.R800004-JLR200. Epub 2008 Feb 27. J Lipid Res. 2008. PMID: 18305313 Free PMC article. Review.
Cited by
-
Purification and Characterization of Mitochondrial Mg2+-Independent Sphingomyelinase from Rat Brain.Mol Cells. 2023 Sep 30;46(9):545-557. doi: 10.14348/molcells.2023.0074. Epub 2023 Jun 12. Mol Cells. 2023. PMID: 37305954 Free PMC article.
-
Sphingolipids in mitochondria.Biochim Biophys Acta Mol Cell Biol Lipids. 2017 Jan;1862(1):56-68. doi: 10.1016/j.bbalip.2016.09.019. Epub 2016 Sep 30. Biochim Biophys Acta Mol Cell Biol Lipids. 2017. PMID: 27697478 Free PMC article. Review.
-
Sphingolipids and lipid rafts: Novel concepts and methods of analysis.Chem Phys Lipids. 2018 Nov;216:114-131. doi: 10.1016/j.chemphyslip.2018.08.003. Epub 2018 Sep 5. Chem Phys Lipids. 2018. PMID: 30194926 Free PMC article. Review.
-
Approaches for probing and evaluating mammalian sphingolipid metabolism.Anal Biochem. 2019 Jun 15;575:70-86. doi: 10.1016/j.ab.2019.03.014. Epub 2019 Mar 24. Anal Biochem. 2019. PMID: 30917945 Free PMC article. Review.
-
Sphingolipids in mitochondria-from function to disease.Front Cell Dev Biol. 2023 Nov 21;11:1302472. doi: 10.3389/fcell.2023.1302472. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 38078003 Free PMC article. Review.
References
-
- Ardail D, Popa I, Alcantara K, Pons A, Zanetta JP, Louisot P, Thomas L, Portoukalian J. Occurrence of ceramides and neutral glycolipids with unusual long-chain base composition in purified rat liver mitochondria. FEBS Lett. 2001;488:160–164. - PubMed
-
- Tserng KY, Griffin R. Quantitation and molecular species determination of diacylglycerols, phosphatidylcholines, ceramides, and sphingomyelins with gas chromatography. Analytical biochemistry. 2003;323:84–93. - PubMed
-
- Yu J, Novgorodov SA, Chudakova D, Zhu H, Bielawska A, Bielawski J, Obeid LM, Kindy MS, Gudz TI. JNK3 signaling pathway activates ceramide synthase leading to mitochondrial dysfunction. J Biol Chem. 2007;282:25940–25949. - PubMed
-
- Futerman AH. Intracellular trafficking of sphingolipids: relationship to biosynthesis. Biochim Biophys Acta. 2006;1758:1885–1892. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
