

Physiological characterization of formyl peptide receptor expressing cells in the mouse vomeronasal organ
- PMID: 25484858
- PMCID: PMC4240171
- DOI: 10.3389/fnana.2014.00134
Physiological characterization of formyl peptide receptor expressing cells in the mouse vomeronasal organ
Abstract
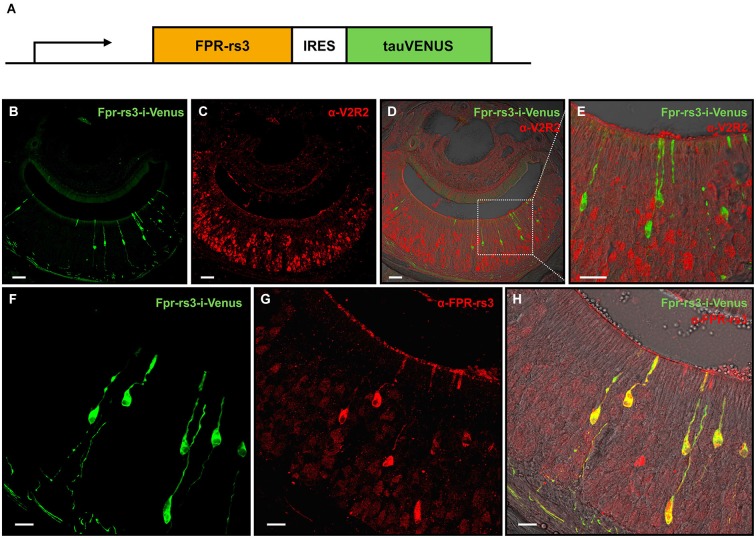
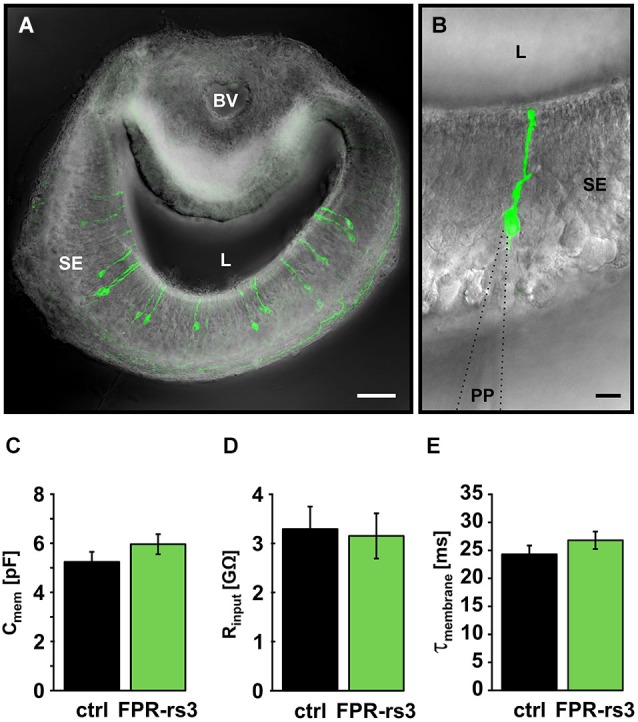
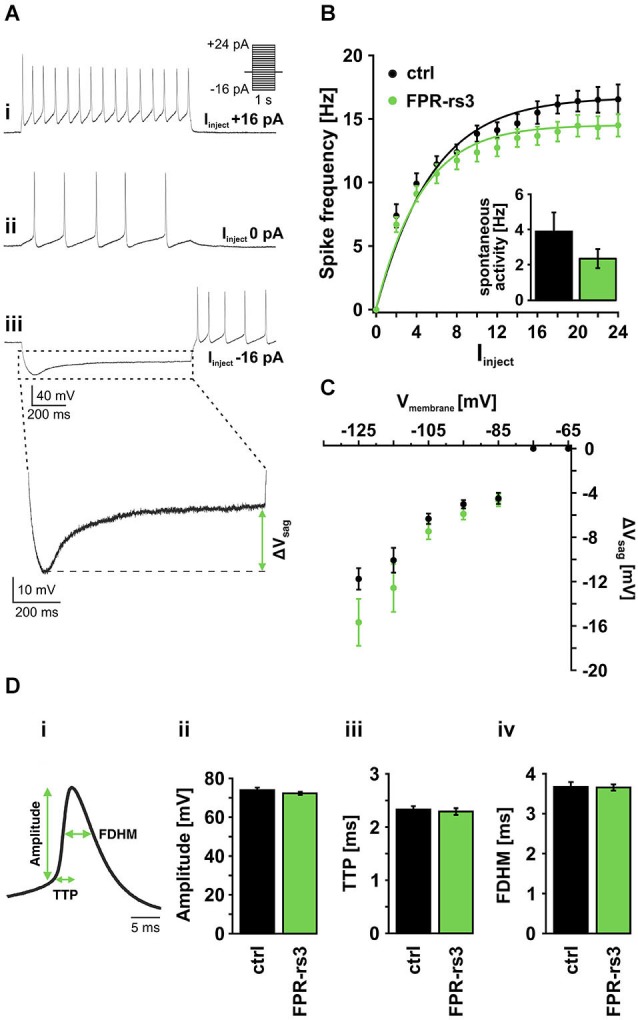
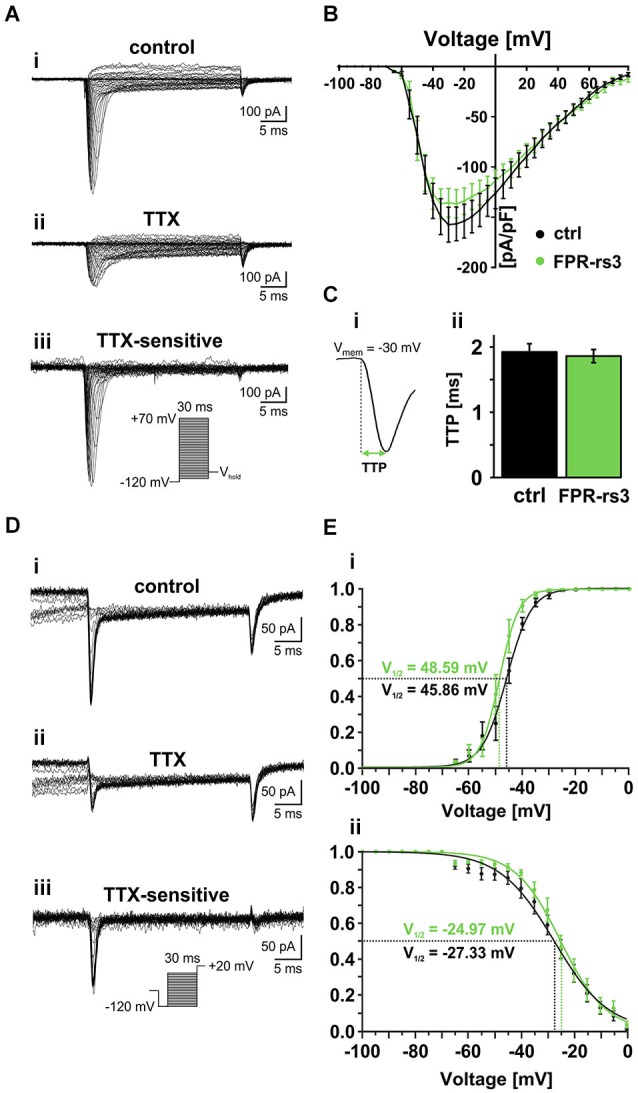
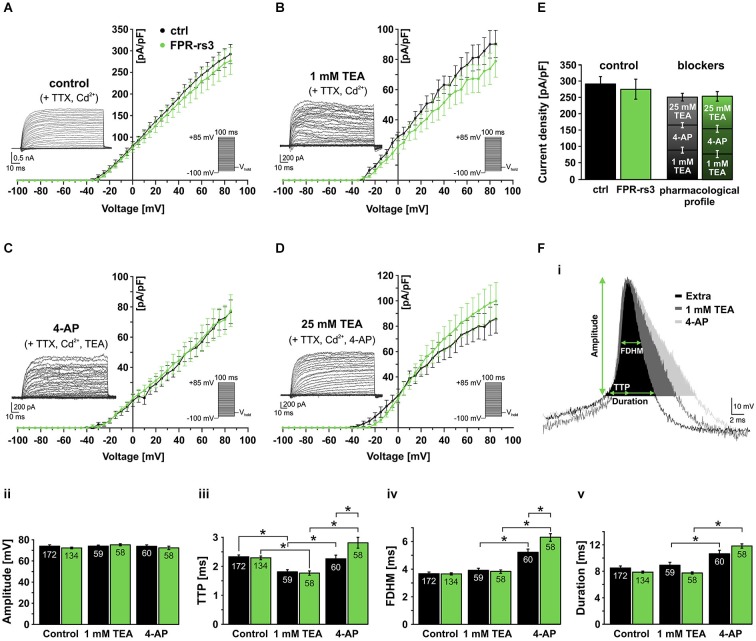
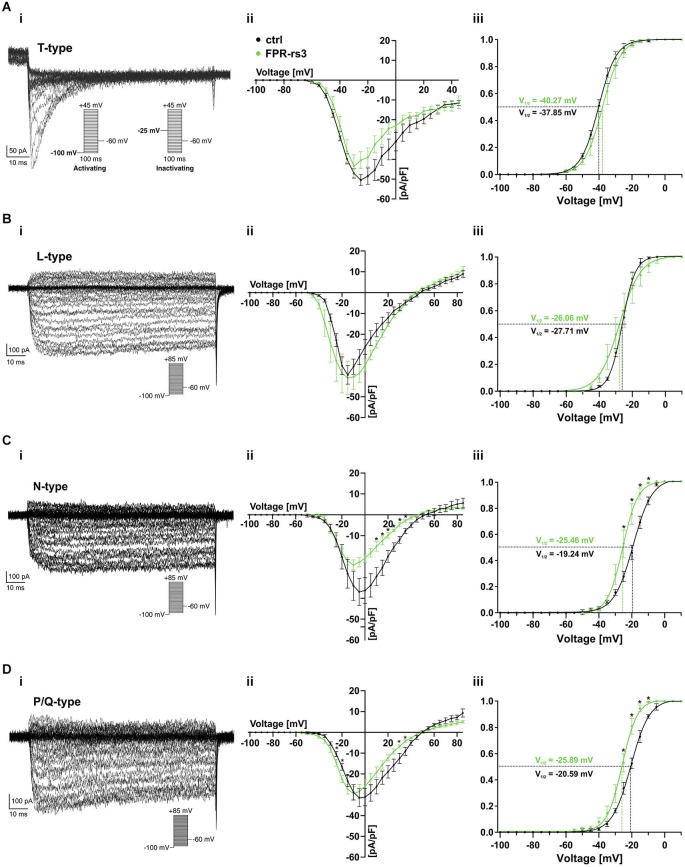
The mouse vomeronasal organ (VNO) is a chemosensory structure that detects both hetero- and conspecific social cues. Based on largely monogenic expression of either type 1 or 2 vomeronasal receptors (V1Rs/V2Rs) or members of the formyl peptide receptor (FPR) family, the vomeronasal sensory epithelium harbors at least three neuronal subpopulations. While various neurophysiological properties of both V1R- and V2R-expressing neurons have been described using genetically engineered mouse models, the basic biophysical characteristics of the more recently identified FPR-expressing vomeronasal neurons have not been studied. Here, we employ a transgenic mouse strain that coexpresses an enhanced variant of yellow fluorescent protein together with FPR-rs3 allowing to identify and analyze FPR-rs3-expressing neurons in acute VNO tissue slices. Single neuron electrophysiological recordings allow comparative characterization of the biophysical properties inherent to a prototypical member of the FPR-expressing subpopulation of VNO neurons. In this study, we provide an in-depth analysis of both passive and active membrane properties, including detailed characterization of several types of voltage-activated conductances and action potential discharge patterns, in fluorescently labeled vs. unmarked vomeronasal neurons. Our results reveal striking similarities in the basic (electro) physiological architecture of both transgene-expressing and non-expressing neurons, confirming the suitability of this genetically engineered mouse model for future studies addressing more specialized issues in vomeronasal FPR neurobiology.
Keywords: VNO; formyl peptide receptor; olfaction; sensory neurons; vomeronasal organ; vomeronasal receptor.
Figures
Similar articles
-
In-depth Physiological Analysis of Defined Cell Populations in Acute Tissue Slices of the Mouse Vomeronasal Organ.J Vis Exp. 2016 Sep 10;(115):54517. doi: 10.3791/54517. J Vis Exp. 2016. PMID: 27684435 Free PMC article.
-
Imaging neuronal responses in slice preparations of vomeronasal organ expressing a genetically encoded calcium sensor.J Vis Exp. 2011 Dec 6;(58):3404. doi: 10.3791/3404. J Vis Exp. 2011. PMID: 22157702 Free PMC article.
-
Convergence of FPR-rs3-expressing neurons in the mouse accessory olfactory bulb.Mol Cell Neurosci. 2013 Sep;56:140-7. doi: 10.1016/j.mcn.2013.04.008. Epub 2013 May 7. Mol Cell Neurosci. 2013. PMID: 23664818
-
Mechanisms underlying pre- and postnatal development of the vomeronasal organ.Cell Mol Life Sci. 2021 Jun;78(12):5069-5082. doi: 10.1007/s00018-021-03829-3. Epub 2021 Apr 19. Cell Mol Life Sci. 2021. PMID: 33871676 Free PMC article. Review.
-
Recent progress in the neurobiology of the vomeronasal organ.Microsc Res Tech. 2002 Aug 1;58(3):228-50. doi: 10.1002/jemt.10094. Microsc Res Tech. 2002. PMID: 12203701 Review.
Cited by
-
The nasal cavity and its olfactory sensory territories.Front Neuroanat. 2015 Mar 18;9:31. doi: 10.3389/fnana.2015.00031. eCollection 2015. Front Neuroanat. 2015. PMID: 25852492 Free PMC article. No abstract available.
-
Ca2+-Activated Ion Channels Exert Opposite Effects in Different Signaling Compartments of Vomeronasal Sensory Neurons.J Neurosci. 2025 Apr 16;45(16):e2134242025. doi: 10.1523/JNEUROSCI.2134-24.2025. J Neurosci. 2025. PMID: 40032527 Free PMC article.
-
In-depth Physiological Analysis of Defined Cell Populations in Acute Tissue Slices of the Mouse Vomeronasal Organ.J Vis Exp. 2016 Sep 10;(115):54517. doi: 10.3791/54517. J Vis Exp. 2016. PMID: 27684435 Free PMC article.
-
Signal Detection and Coding in the Accessory Olfactory System.Chem Senses. 2018 Nov 1;43(9):667-695. doi: 10.1093/chemse/bjy061. Chem Senses. 2018. PMID: 30256909 Free PMC article. Review.
-
The Effects of Essential Oils and Terpenes in Relation to Their Routes of Intake and Application.Int J Mol Sci. 2020 Feb 25;21(5):1558. doi: 10.3390/ijms21051558. Int J Mol Sci. 2020. PMID: 32106479 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources