

Extracellular ATP activates MAPK and ROS signaling during injury response in the fungus Trichoderma atroviride
- PMID: 25484887
- PMCID: PMC4240048
- DOI: 10.3389/fpls.2014.00659
Extracellular ATP activates MAPK and ROS signaling during injury response in the fungus Trichoderma atroviride
Abstract
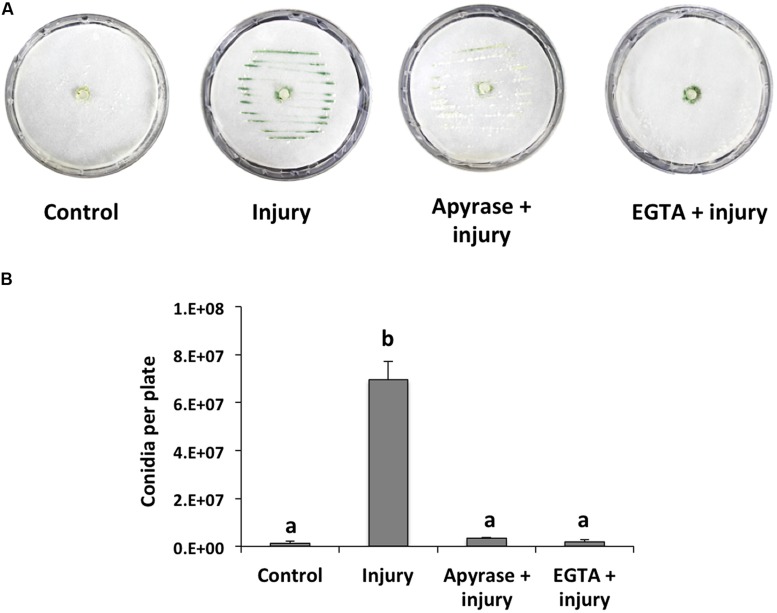
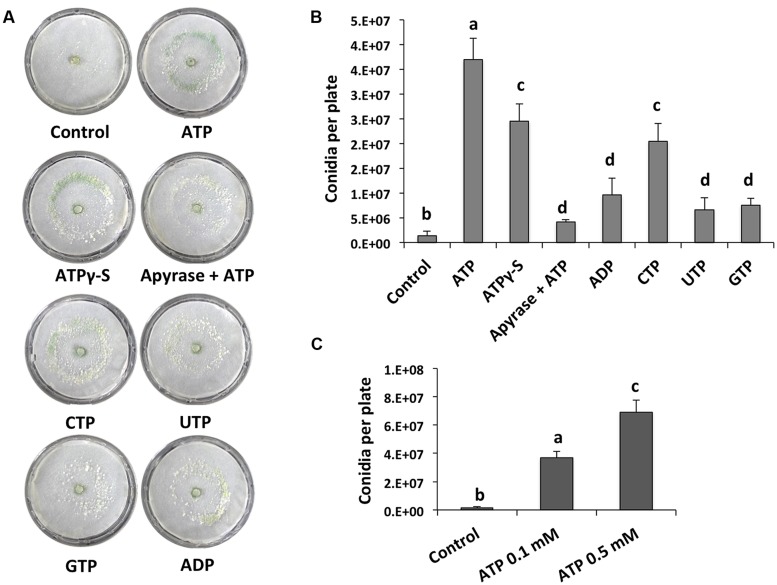
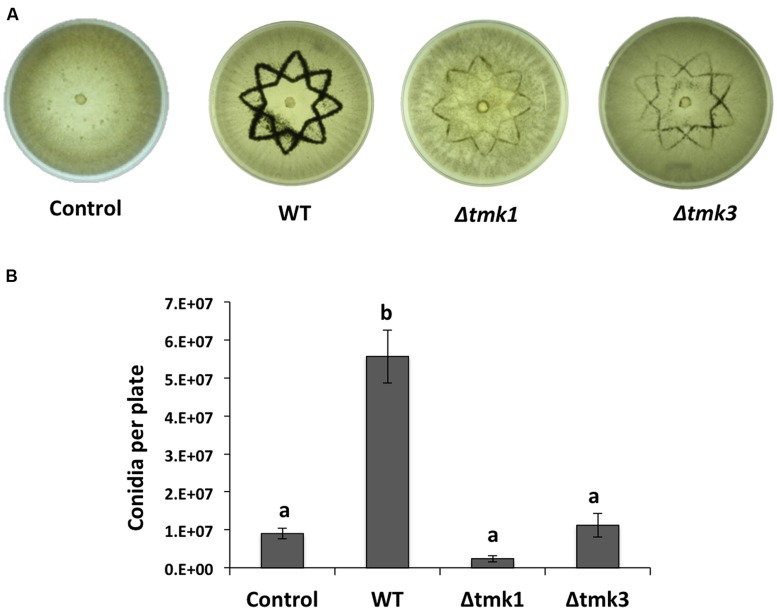
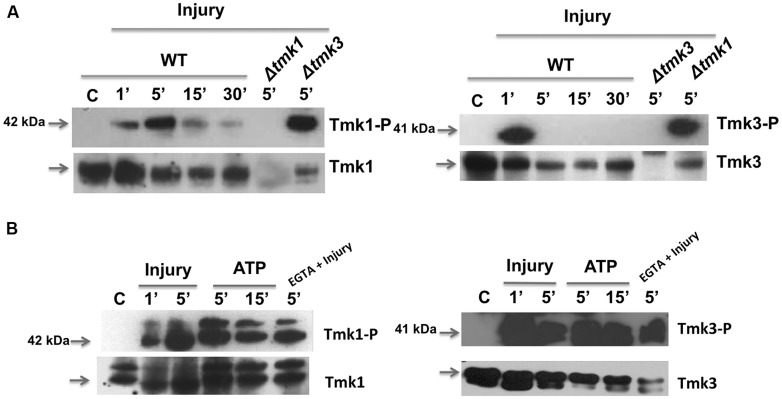
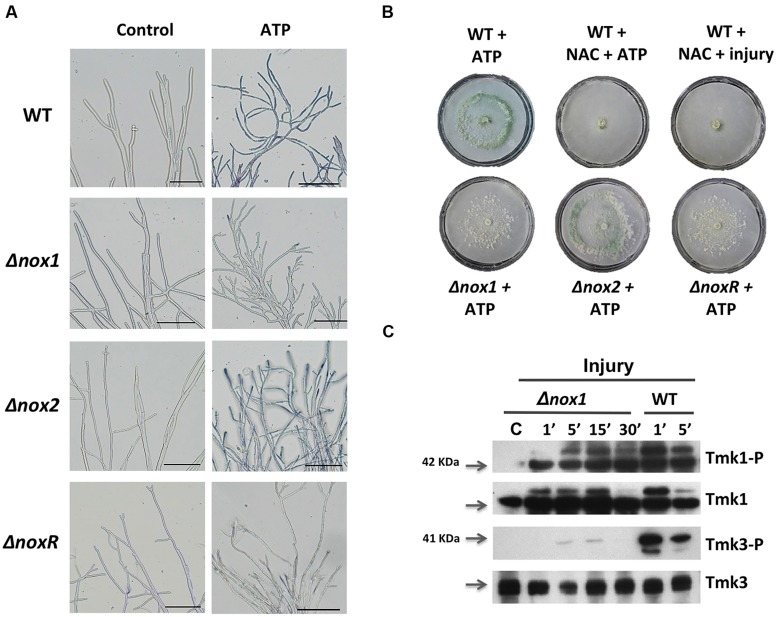
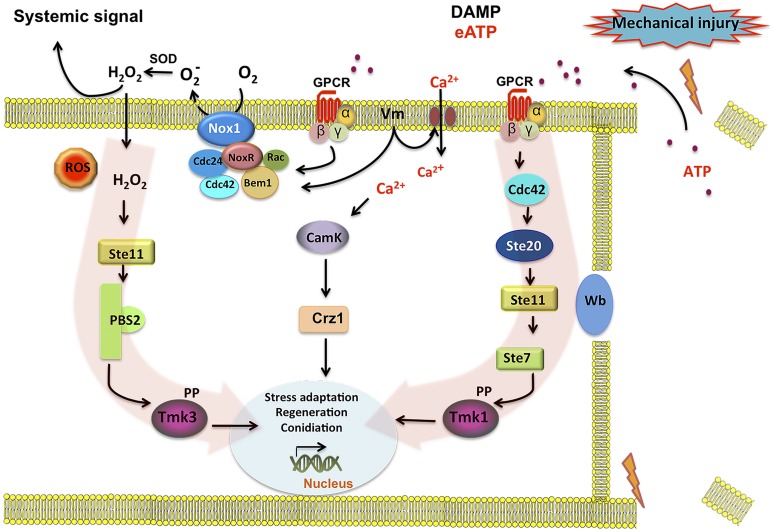
The response to mechanical damage is crucial for the survival of multicellular organisms, enabling their adaptation to hostile environments. Trichoderma atroviride, a filamentous fungus of great importance in the biological control of plant diseases, responds to mechanical damage by activating regenerative processes and asexual reproduction (conidiation). During this response, reactive oxygen species (ROS) are produced by the NADPH oxidase complex. To understand the underlying early signaling events, we evaluated molecules such as extracellular ATP (eATP) and Ca(2+) that are known to trigger wound-induced responses in plants and animals. Concretely, we investigated the activation of mitogen-activated protein kinase (MAPK) pathways by eATP, Ca(2+), and ROS. Indeed, application of exogenous ATP and Ca(2+) triggered conidiation. Furthermore, eATP promoted the Nox1-dependent production of ROS and activated a MAPK pathway. Mutants in the MAPK-encoding genes tmk1 and tmk3 were affected in wound-induced conidiation, and phosphorylation of both Tmk1 and Tmk3 was triggered by eATP. We conclude that in this fungus, eATP acts as a damage-associated molecular pattern (DAMP). Our data indicate the existence of an eATP receptor and suggest that in fungi, eATP triggers pathways that converge to regulate asexual reproduction genes that are required for injury-induced conidiation. By contrast, Ca(2+) is more likely to act as a downstream second messenger. The early steps of mechanical damage response in T. atroviride share conserved elements with those known from plants and animals.
Keywords: calcium; conidiation; extracellular ATP (eATP); injury response; mitogen-activated protein kinase (MAPK); reactive oxygen species (ROS).
Figures
References
-
- Abbracchio M. P., Burnstock G., Boeynaems J.-M., Barnard E. A., Boyer J. L., Kennedy C., et al. (2006). International union of pharmacology LVIII: update on the P2Y G protein-coupled nucleotide receptors: from molecular mechanisms and pathophysiology to therapy. Pharmacol. Rev. 58 281–341 10.1124/pr.58.3.3 - DOI - PMC - PubMed
-
- Beneloujaephajri E., Costa A., L’haridon F., Métraux J.-P., Binda M. (2013). Production of reactive oxygen species and wound-induced resistance in Arabidopsis thaliana against Botrytis cinerea are preceded and depend on a burst of calcium. BMC Plant Biol. 13:160 10.1186/1471-2229-13-160 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous