

Role of chaperones and ATP synthase in DNA gyrase reactivation in Escherichia coli stationary-phase cells after nutrient addition
- PMID: 25485196
- PMCID: PMC4230433
- DOI: 10.1186/2193-1801-3-656
Role of chaperones and ATP synthase in DNA gyrase reactivation in Escherichia coli stationary-phase cells after nutrient addition
Abstract
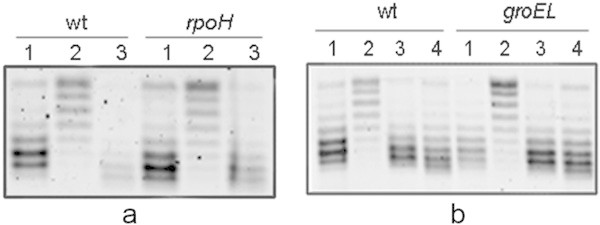
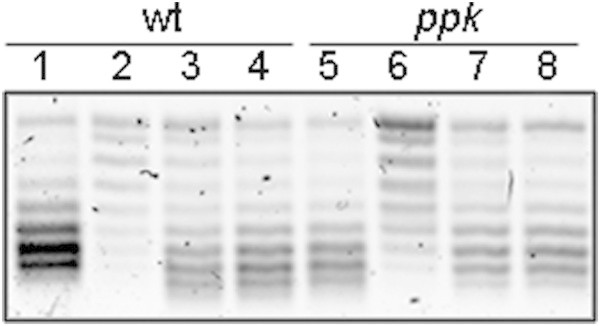
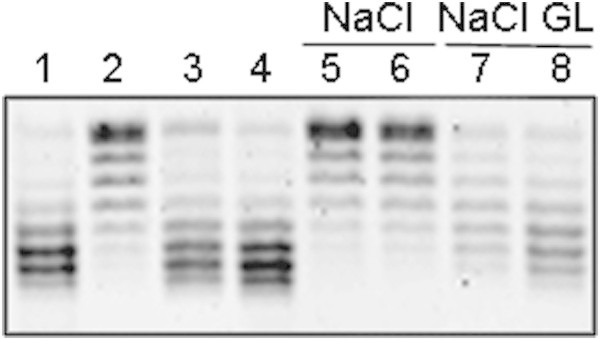
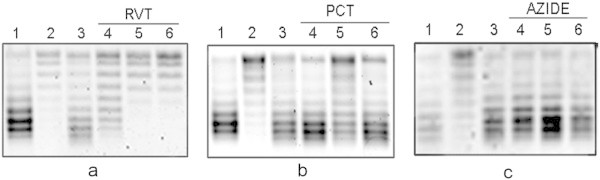
Escherichia coli stationary-phase (SP) cells contain relaxed DNA molecules and recover DNA supercoiling once nutrients become available. In these cells, the reactivation of DNA gyrase, which is a DNA topoisomerase type IIA enzyme, is responsible for the recovery of DNA supercoiling. The results presented in this study show that DNA gyrase reactivation does not require cellular chaperones or polyphosphate. Glucose addition to SP cells induced a slow recovery of DNA supercoiling, whereas resveratrol, which is an inhibitor of ATP synthase, inhibited the enzyme reactivation. These results suggest that DNA gyrase, which is an ATP-dependent enzyme, remains soluble in SP cells, and that its reactivation occurs primarily due to a rapid increase in the cellular ATP concentration.
Keywords: ATP synthase; DNA gyrase; Escherichia coli; Stationary phase.
Figures
References
-
- Ahn K, Kornberg A. Polyphosphate kinase from Escherichia coli. Purification and demonstration of a phosphoenzyme intermediate. J Biol Chem. 1990;265(20):11734–11739. - PubMed
-
- Bochner BR, Ames BN. Complete analysis of cellular nucleotides by two dimensional thin-layer chromatography. J Biol Chem. 1982;257:9759–9769. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
